Czym jest kręgosłup piersiowy ssaków. Budowa zewnętrzna ssaków, szkielet i mięśnie na przykładzie psa domowego
Kręgosłup składa się z odcinka szyjnego, piersiowego, lędźwiowego, krzyżowego i ogonowego. Jego charakterystyczna cecha- platylialny (o płaskich powierzchniach) kształt kręgów, pomiędzy którymi znajdują się chrząstki krążki międzykręgowe... Górne łuki są dobrze zaznaczone. W odcinku szyjnym znajduje się siedem kręgów, których długość zależy od długości szyi; tylko w manatach i leniwcach - Choloepus hof-fmani jest ich 6, aw leniwcu - Bradypus 8-10. Kręgi szyjne są bardzo długie u żyraf i bardzo krótkie u waleni, które nie mają odcinka szyjnego. Żebra tworzące klatkę piersiową są przymocowane do kręgów odcinka piersiowego. Zamykający go mostek jest płaski i tylko przy nietoperze a gatunki kopiące nory z potężnymi kończynami przednimi (na przykład pieprzyki) mają mały grzebień (kil) służący jako punkt mocowania mięśni piersiowych. W odcinku piersiowym znajduje się 9-24 (zwykle 12-15) kręgów, ostatnie 2-5 kręgów piersiowych - fałszywe żebra - nie sięgające mostka., W odcinku lędźwiowym od 2 do 9 kręgów; szczątkowe żebra łączą się z ich dużymi procesami poprzecznymi. Region sakralny składa się z 4-10 zrośniętych kręgów, z których tylko dwa pierwsze są rzeczywiście sakralne, a pozostałe są ogonowe. Liczba wolnych kręgów ogonowych waha się od 3 (w gibonie) do 49 u dinozaura z długim ogonem.
Stopień ruchomości poszczególnych kręgów jest różny. U małych biegających i wspinających się zwierząt jest duży, na całej długości kręgosłupa, więc ich ciało może się zginać różne kierunki a nawet zwinąć się w kulkę. Kręgi w odcinku piersiowym i lędźwiowym są mniej ruchliwe u dużych, szybko poruszających się zwierząt. U ssaków podróżujących dalej tylne nogi ah (kangury, skoczki, skoczki), największe kręgi znajdują się u nasady ogona i kości krzyżowej, a dalej ich rozmiar stopniowo maleje. Natomiast u zwierząt kopytnych kręgi, a zwłaszcza ich wyrostki kolczyste, są większe w przedniej części odcinka klatki piersiowej, gdzie są przymocowane do nich potężne mięśnie szyi i częściowo kończyn przednich (
Zewnętrzna budowa psa
Pies był pierwszym zwierzakiem. Człowiek oswoił ją w starożytności. Pies pomagał prymitywnemu człowiekowi podczas polowania, pilnował jego domu. Znane są obecnie psy służbowe, myśliwskie i ozdobne, a także kundle (kundle). Psy pomagają w pełnieniu służby ochrony granic naszej Ojczyzny, a w okresie Wielkim Wojna Ojczyźniana szukał rannych, pomagał w rozpoznaniu i komunikacji. Na psach naukowcy prowadzą badania żywotnej aktywności organizmu. Pies przed człowiekiem odwiedzona przestrzeń.
Ciało psa, takiego jak pasterz czy husky, jest smukłe i muskularne. Jego kończyny znajdują się nie po bokach ciała, jak u gadów, ale pod ciałem, dzięki czemu ciało zwierzęcia nie dotyka ziemi. Pies chodzi opierając się na palcach z mocnymi pazurami. Elastyczna szyja zapewnia doskonałą mobilność głowy. Pysk zwierzęcia jest ograniczony ruchomymi wargami – górnymi i dolnymi. Nad Górna warga jest nos z parą zewnętrznych otworów nosowych - nozdrza. Oczy mają dobrze rozwinięte powieki. Mrugająca błona (trzecia powieka) u psa, jak u wszystkich ssaków, jest słabo rozwinięta. Ze wszystkich zwierząt tylko ssaki mają ucho zewnętrzne - Małżowina uszna... Ucho zewnętrzne psa jest duże i ruchome.
Pokrowce na ssaki
Skóra ssaków jest mocna i elastyczna, u większości zwierząt znajdują się w niej nasady włosów, które tworzą charakterystyczną dla tej klasy kręgowców linia włosów ... Rozróżnij grube i długie włosy — kręgosłup i krótsze, bardziej miękkie - pod podłogą, lub podkład... Gruba i wytrzymała markiza chroni podszerstek i skórę przed uszkodzeniami. Podszerstek, który zatrzymuje dużo powietrza, dobrze zatrzymuje ciepłotę ciała. Oprócz markizy i podszycia rozwijają się zwierzęta duże włosy- narządy dotyku.
Sierść ssaków, podobnie jak pióra ptaków i łuski gadów, składa się z materii rogowej. Dwa razy w roku pies, podobnie jak wiele innych ssaków, linieje – część jego sierści wypada i jest zastępowana nowymi. Podstawa włosów znajduje się w specjalnej torbie na włosy, w której otwierają się kanaliki znajdujące się w pobliżu gruczoły łojowe... Ich wydzieliny natłuszczają skórę i włosy, dzięki czemu są elastyczne i nie zwilżane wodą.
Wzór: struktura skóry i sierści ssaka
W skórze większości zwierząt znajdują się i gruczoły potowe ... Pot odparowuje z powierzchni ciała i chłodzi je. Wraz z potem z organizmu wydalany jest również nadmiar soli i mocznika; w ten sposób gruczoły potowe odgrywają rolę dodatkowe ciała rozładować się.
Pies gruczoły potowe mało i chłodzenie ciała osiąga się przy wzmożonym oddychaniu.
Na końcach palców ssaków znajdują się zrogowaciałe pazury, pazury lub kopyta. Czasami zrogowaciałe formacje rozwijają się również na głowie (rogi u nosorożców, antylop, bydła itp.) lub na ogonie (na przykład zrogowaciałe łuski u szczurów).
Szkielet ssaka
Szkielet ssaków składa się z tych samych fragmentów, co u innych kręgowców. Czaszka zwierząt wyróżnia się większą czaszką, co jest związane z duży rozmiar mózg. Dla ssaków bardzo charakterystyczna jest obecność 7 kręgów szyjnych. Zarówno żyrafa o długiej szyi, jak i wieloryby mają taką samą liczbę kręgów szyjnych. Kręgi piersiowe (zwykle 12-15) wraz z żebrami i mostkiem tworzą solidną klatkę piersiową. Masywne kręgi lędźwiowy ruchomo połączone ze sobą. W tej sekcji ciało może się zginać i wyginać. Liczba kręgów lędźwiowych różni się w różne rodzaje(2-9), pies ma 6. Kręgosłup krzyżowy (3-4 kręgi) łączy się z kośćmi miednicy. Liczba kręgów ogonowych (od trzech do kilkudziesięciu) zależy od długości ogona.

Rysunek: Szkielet ssaków na przykładzie psa

Wzór: czaszka psa
Pas przednich kończyn ssaki składają się z dwóch łopatek z przyczepionymi do nich kruczymi kośćmi i dwóch obojczyków. Obojczyki psa nie są rozwinięte. Pas na tylną kończynę- miednica - utworzona przez trzy pary kości miednicy... Szkielety kończyn u różnych gatunków ssaków i gadów są podobne, jednak szczegóły ich budowy u różnych gatunków są różne i zależą od warunków życia zwierzęcia.
Mięśnie ssaków
Większość ssaków, w tym pies, ma szczególnie rozwinięte mięśnie grzbietu, kończyn i ich pasów. Pies może biegać dużymi skokami, zginając i prostując ciało, na przemian odpychając się od ziemi przednimi lub tylnymi łapami. Silne mięśnie poruszają dolną szczęką. Zaciskając szczęki, uzbrojony w zęby, pies mocno trzyma zdobycz.

Wzór: Mięśnie psa poruszającego dolną szczęką
Obręczy barkowej ssaki są uproszczone, składają się z dużego łopatki o wyrostku krukowatym, i obojczyk(ryc. 80, 81). W embrionalnym stadium rozwoju oraz w przypadku dorosłych stekowców krukowaty jest niezależną kością.
Łopatka znajduje się na górze Klatka piersiowa, ma trójkątny kształt i wysoki wyrost (grzebień) z akromia proces mocowania mięśni. W miejscu wyrostka kruczego i wyrostka barkowego łopatki znajduje się dołek panewkowy do mocowania kończyny przedniej.
Obojczyk w kształcie pręta służy do połączenia barku i mostka. Obojczyk jest najbardziej rozwinięty u zwierząt wykonujących kończyny przednie. złożone ruchy(nietoperze, krety, naczelne, gryzonie itp.). U wielu gatunków, które charakteryzują się jednostajnymi ruchami (kopytne, psy), nie ma obojczyka. Ogólnie rzecz biorąc, obręcz kończyn przednich jest połączona ze szkieletem osiowym poprzez mięśnie i więzadła.
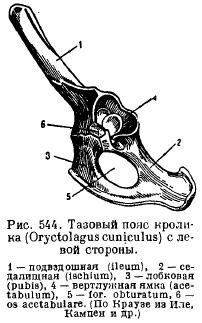
Obręcz miednicy (rys. 82 A) składa się z sparowanych bezimienne kości przegubowy biodrowa, kulszowa i łonowa kości. Miednica zamknięty typ i przez biodro jest mocno połączony z sakralny kręgosłup. Kiedy elementy miednicy rosną razem, związek publiczny i uformowany panewka do mocowania kończyn tylnych. W dolnej części kości pierścieniowej znajduje się otwór obturatora.
Sparowany szkielet kończyn
Kończyny ssaków, podobnie jak innych kręgowców lądowych, są typu pięciopalczastego i reprezentują trójczłonową dźwignię, której wszystkie części są ze sobą ruchomo połączone.
Przednia kończyna zawiera ramię, przedramię i dłoń(rys. 81). Kość ramienna dobrze rozwinięta, posiada zaokrągloną główkę do mocowania do obręczy kończyny. Przedramię zrób dwie kości - promieniowy i łokciowy mający olecranon na górze. Na styku barku i przedramienia znajduje się staw łokciowy.
Szczotka obejmuje trzy działy - nadgarstek, śródręcze i paliczki. Nadgarstek składa się z 8-10 elementów ułożonych w trzech rzędach ... Pęcina zawiera pięć kości (w zależności od liczby palców). Każdy palec ma trzy paliczki, z wyjątkiem pierwszego palca, który ma dwa paliczki. W przeciwieństwie do artykulacji międzynadgarstkowej gadów i ptaków, telefon komórkowy wspólny u ssaków zlokalizowanych między przedramieniem a dłonią(jak płazy).
Kończyny tylnej ssaki mają również trzy działy - uda, podudzie i stopa(Rys. 82 B). Kość udowa masywny i włożony zaokrągloną głową w panewkę obręczy miednicy. Goleń zawiera dwie kości - piszczel i piszczel. Większa piszczel jest wewnętrznym elementem kości piszczelowej, a strzałkowa jest jej zewnętrzną częścią.
Pomiędzy udem a podudziem znajduje się wspólny pokryty nakolannik które wyewoluowały z ich skostniałych ścięgien mięśni. Powierzchnie kości stawowych pokryte są formacjami chrzęstnymi. Stawu skokowego ssaków znajduje się między dolnymi końcami kości goleni a proksymalną częścią stopy.

Ryż. 82. Obręcz miedniczna (A) i tylna kończyna (B) lisa:
1 – biodro, 2 - kulsz, 3 - kość łonowa, 4 - panewka, 5 - kość udowa, 6 - rzepka, 7 - piszczel, 8 - strzałka, 9 - stęp, 10 - śródstopie, 11 - paliczki palców, 12 - kość skokowa, 13 - pięta
Elementy stępu stopy są ułożone w trzech rzędach i tworzą narośl - kości piętowej typowy dla ssaków. Śródstopie składa się z pięciu kości, do których przymocowane są paliczki palców. Podobnie jak w kończynach przednich, w każdym palcu znajdują się trzy z nich, z wyjątkiem pierwszego (wewnętrznego), który ma tylko dwa paliczki.
Dla ssaków charakterystyczne jest specjalne ustawienie kończyn względem ciała: znajdują się one pod ciałem, co przyczynia się do rozwoju większej manewrowości ruchów. W tym przypadku staw łokciowy jest skierowany do tyłu i staw kolanowy przeciwnie, jest zorientowany do przodu.
W związku z różnorodnością gatunkową ssaków, ich trybem życia i sposobami poruszania się, względna długość kończyn, stosunek elementów składowych, konfiguracja kości, liczba palców itp. znacznie się różnią. u nietoperzy długość kończyny przedniej, zwłaszcza paliczków palców, służąca do mocowania błony powietrznej.
Przeciwnie, przednia kończyna pieprzyka jest skrócona, część dłoniowa poszerzona, a szósty palec u nogi jest dobrze rozwinięty. U wielorybów ręka jest ostro wydłużona ze względu na wzrost liczby paliczków palców, w wyniku czego nabyła kończyna wielkie podobieństwo z płetwą z krzyżowej ryby. Palce zwierząt o dużej biomasie i szybkim wybiegu (koniowate, parzystokopytne) zostały znacznie zredukowane i zmodyfikowane.
Jedną z cech szkieletu ssaków jest wielki rozwój chrząstka u młodych zwierząt i jej długotrwała konserwacja w okresie rozwoju poembrionalnego. Wynika to ze specyfiki wzrostu chrząstki i kości w związku ze wzrostem samego zwierzęcia. To chrząstka rośnie całą swoją masą (śródmiąższowo), wzrost następuje szybko i proporcje części mogą być zachowane, natomiast kość rośnie wolniej i tylko z powierzchni, tak że części szkieletu, które pojawiają się zawsze pozostają w tej samej odległości, a zachowanie proporcji części wraz ze wzrostem kości okazuje się niemożliwe. Dlatego młode zwierzęta zawsze zawierają dużo chrząstki, a zwłaszcza w miejscach, w których następuje wzrost. Więc, długie kości kończyny zawsze składają się z części środkowej - trzonu, który przede wszystkim kostnieje, oraz końców, które są zachowane przez długi czas przez chrząstkę - szyszynkę. Kostnienie trzonu zaczyna się przede wszystkim od obwodu - okołochrzęstnego, czyli z powodu komórek pochodzących z tkanki łącznej pokrywającej chrząstkę - z ochrzęstnej; następnie łączy się z nią kostnienie wewnętrzne chrząstki - kostnienie śródchrzęstne; dopiero później, gdy w trzonie kości w wyniku rozpadu kości pojawią się ubytki szpiku kostnego, szyszynka zaczyna kostnieć endochondrycznie; między trzonem a nasadą jednak chrząstka pozostaje przez długi czas, za pomocą której kość rośnie na długość z zachowaniem proporcji (ryc. 530).
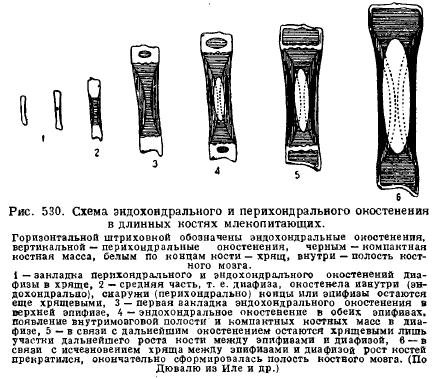
Kręgosłup ssaki charakteryzują się, w przeciwieństwie do zauropsydy, brakiem wolnych stawów między kręgami, z wyjątkiem pierwszego i drugiego kręgu. Międzykręgowe elastyczne krążki tkanki łącznej leżą między trzonami kręgów. Na ogół skutkuje to mniejszą elastycznością kręgosłupa, co wynika z bardziej wyspecjalizowanego ruchu ssaków przy pomocy kończyn.
Kostnienie trzonów kręgów następuje przez szyszynkę chrzęstną. Ciała mają powierzchnie płaskie lub lekko wypukłe, u większości współczesnych zwierząt kopytnych są nieco opistoceeliczne. Na kręgach wyrostki kolczyste są dobrze rozwinięte, zwłaszcza w odcinku piersiowym, a także wyrostki poprzeczne, które znajdują się na łukach; u podstawy tego ostatniego znajdują się procesy stawowe.

Liczba kręgów szyjnych wynosi zwykle 7 (ryc. 531), niezależnie od długości szyi, więc długość szyi, w przeciwieństwie do ptaków, zależy wyłącznie od długości kręgów. Tylko manat (Manatus) i jeden gatunek leniwca (Choloepus hoffmani) mają 6 kręgów szyjnych, podczas gdy leniwce trójpalczaste (Bradypus) mają 9. Szyja waleni, która prawie zniknęła z powodu wodnego trybu życia, nadal zawiera 7 płaskich , częściowo zlewające się kręgi, podczas gdy w szyi żyrafy te 7 kręgów jest bardzo długich. Pierwsze dwa kręgi szyjne są zmodyfikowane w atlas i epistrofię, połączone prawdziwymi stawami, a u stekowców proces odontoidu nadal funkcjonuje w postaci specjalnej kości i nie zlał się, jak u innych ssaków, z ciałem epistrofia. Wraz z kręgami rosną szczątkowe żebra odcinka szyjnego, w wyniku czego powstają perforowane procesy poprzeczne. Częściowe lub całkowite zespolenie kręgów szyjnych występuje u waleni; u skoczek pustynnych wszystkie kręgi szyjne rosną razem z wyjątkiem siódmego, w krecie torbacza (Notoryctes) łączą się od 2 do 6, w pieprzu - od 3 do 4.
Liczba kręgów piersiowych, które charakteryzują przyczepione do nich żebra, waha się od 9 [u jednego waleni (Hyperoodon) i pancernika (Tatusia)] do 25 [u leniwca], ale zwykle jest ich 12-13. Procesy na kręgach piersiowych są dobrze rozwinięte: kolczaste (proc. Spinosus), poprzeczne lub diapofizy (proc. Transversus), stawowe lub zygapofizy (proc. Articularis); żebra są połączone głowami z korpusem jednego lub dwóch sąsiednich kręgów oraz guzkami z wyrostkami poprzecznymi. Składają się z dwóch części: żebra kostnego - proksymalnego i chrzęstno - dystalnego. Tylko żebra przednich kręgów piersiowych docierają do mostka, tylne żebra kończą się swobodnie (żebra fałszywe). Kręgi lędźwiowe, które charakteryzują się brakiem żeber wchodzących tu w skład wyrostków poprzecznych, u ssaków od 2 do 9, zwykle 6-7.
Kręgi krzyżowe ssaków to te kręgi, które zrosły się razem i są połączone przegubowo kość biodrowa... Takich kręgów jest 1-10, ale tylko 1-2 z nich to prawdziwe kręgi krzyżowe, czyli takie, w których połączenie z miednicą następuje poprzez niezależne skostnienia odpowiadające żebrom krzyżowym; pozostałe kręgi krzyżowe są fałszywe, to znaczy kręgi ogonowe przymocowane do kości krzyżowej.
Liczba kręgów zawartych w regionie ogonowym jest bardzo zróżnicowana: od 3 ( nietoperze, gibon) do 49 [jaszczurka (Manis macrura)].
Mostek(mostek) ssaków rozwija się z połączenia brzusznych końców żeber, które następnie tworzą jedną niesparowaną płytkę, składającą się z kilku działów (ryc. 532).
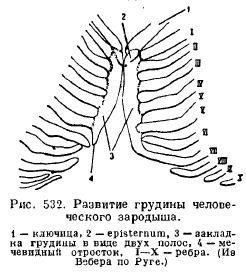
Połączony z obojczykami (jeśli występują) i z pierwszą parą żeber, przedłużony mostek przedni nazywany jest uchwytem (praesternum, manubrium); następnie oddział zwany ciałem (mesosternum, corpus), często dzielący się na szereg kości, kolejno umieszczonych między kolejnymi parami żeber; jeszcze dalej znajduje się tak zwany proces wyrostka mieczykowatego (xiphisternum). Nie wszystkie żebra są połączone z mostkiem, mniej lub bardziej znacząca liczba fałszywych żeber kończy się swobodnie (ryc. 533).

U ssaków kloakalnych (Monotremata), ze względu na obecność kruków, których nie ma u wszystkich innych ssaków, przed uchwytem mostka układana jest chrząstka mostka (prosternum), co jest związane z mostkiem. wyrostka kruczego, który jest następnie wypierany przez rozwijającą się kość - episternum (episternmn), co zbliża chrząstkę kloakalną do gadów.
Wiosłować ssaki różnią się wieloma cechami od czaszki gadów. Jednak te cechy nie pojawiły się natychmiast, ale poprzez stopniową zmianę, którą można spotkać badając czaszki kopalnych jaszczurek (Theromorpha), zwłaszcza z rzędu zwierząt o zębach zwierzęcych (Theriodontia). Czaszka ssaków - czaszka tropikalna, ale ze skróconą częścią międzyoczodołową, przez co wygląda jak płaskopodstawna, z dużą czaszką, z wyraźną tendencją do zrastania się poszczególnych kości ze sobą,
Umiejscowienie kości w czaszce ssaków można ocenić na podstawie załączonego diagramu (ryc. 534).
Na bocznych kościach potylicznych, na zewnątrz od guzków stawowych, zwykle występuje wyrostek potyliczny (str. Paroccipitalis), który jest szczególnie dobrze rozwinięty u gryzoni i wielu kopytnych. Przed guzkami stawowymi zwykle znajduje się otwór na zewnątrz nerwu gnykowego (XII) (np. Condyloideum anterius).
W chrząstce torebki słuchowej znajduje się kilka skostnień, które jednak bardzo wcześnie łączą się w dwie kości: leżącą z przodu - kamienistą (petrosum) i leżącą na zewnątrz i za - wyrostek sutkowaty (mastoideum); obie te kości, połączone razem, dają wyrostek sutkowaty lub po prostu kość kamienną (petromastoideum, petrosum lub okostną), obejmującą wewnętrzny błędnik słuchowy. Granice okostnej w tylnej części z główną i boczną kość potyliczną, w przedniej części z kośćmi klinowymi i skrzydłowo-klinowymi; ponieważ naczynia krwionośne i nerwy przechodzą między tymi kośćmi a kośćmi kamienistymi, pozostają tu znaczne luki: z tyłu - rozdarty otwór tylny (dla lacerum posterius) lub otwór szyjny (dla jugulare) dla przejścia nerwu językowo-gardłowego (IX), gałęzie nerwu błędnego ( X) i nerwu dodatkowego (XI), a także żyły szyjnej wewnętrznej, a z przodu - przedni otwór rany (w przypadku Lacerum anterius), przez który tętnica szyjna wewnętrzna przechodzi do czaszki . Niekiedy połączenie kości skalistej z czaszką jest tak słabe, że można ją łatwo oddzielić wraz z przylegającą do niej kością bębenkową, a nawet przy maceracji czaszki całkowicie wypada, jak to ma miejsce u waleni.
Poza obszarem ucha tworzą się dwie kości pokrywające. Jeden z nich, którego pochodzenie wciąż nie jest dostatecznie wyjaśnione, rozwija się w okolicy przewodu słuchowego zewnętrznego w postaci półkolistej małej kości. Jest to kość bębenkowa (tympanicum), która prawdopodobnie jest homologiem kości kątowej (angulare) żuchwa gady, które przeniosły się do okolicy przyusznej wraz z innymi kośćmi żuchwy z powodu zmian w artykulacji szczęki. Zlokalizowany początkowo w postaci kości półkolistej lub pierścieniowej w okolicy przewodu słuchowego, często rozrasta się i tworzy, zwykle wraz z kością kamienistą, a czasem łuszczącą się, przewód słuchowy zewnętrzny (meatus auditius externus). Błona bębenkowa rozciąga się na kości bębenkowej. Głębiej do wewnątrz kość bębenkowa może być również zaangażowana w tworzenie ścianek ucha środkowego. Często kość bębenkowa na spodzie obszaru słuchowego pęcznieje w cienkościenną bańkę (bulla ossea).
Drugą kością pochodzenia skórnego, która bierze udział w tworzeniu szkieletu okolicy ucha, jest kość płaskonabłonkowa (płaskonabłonek); uzyskał wyjątkowy rozwój i znaczenie u ssaków w związku z przywiązaniem do niego żuchwy. U małp i ludzi wszystkie te kości regionu ucha rosną razem, tworząc złożoną część skroniową; kość płaskonabłonkowa stanowi tutaj tylko część, a mianowicie łuski (squama temporalis) kość skroniowa, podczas gdy kość bębenkowa jest częścią bębenkową (pars tympanicum), a przyuszne kości chrzęstne same tworzą części kamieniste (pars petrosa) i wyrostka sutkowatego (pars mastoidea). Stopień rozwoju kości płaskonabłonkowej i jej udział w tworzeniu ścian puszki mózgowej i przewodu słuchowego są bardzo różne, ale kość płaskonabłonkowa ma zawsze powierzchnię stawową (cavitas glenoidea) dla stawu żuchwy. Tak więc ssaki, w przeciwieństwie do wszystkich innych kręgowców, mają bardzo szczególne, oryginalne przywiązanie żuchwy: nie do kości kwadratowej, ale do płaskonabłonkowej kości czaszki. Powierzchnia stawowa żuchwy leży na podstawie rozciągającej się od łusek na zewnątrz i do przodu proces jarzmowy(proc. zygomaticus). U stekowców nad powierzchnią stawową jest on penetrowany przez kanał skroniowy (can.temporalis), co jest szczególnie wyraźne w dziobaku. Za dołem stawowym znajduje się proces (proc. Postglenoideus).
Przed częścią słuchową podstawę czaszki tworzy główna kość klinowa (basisphenoidale), na wewnętrzna powierzchnia która ma mniej lub bardziej znaczne zagłębienie – tzw. siodło tureckie (sella turtica), w którym znajduje się przysadka mózgowa. Po bokach głównej kości klinowej leżą, tworząc dolne części przedniej ściany czaszki, kości skrzydłowo-klinowe (alisphenoidea), zwykle zrośnięte z nią w jedną kość, tworząc duże skrzydła (alae magnae) tej kości. Kości skrzydłowo-klinowe są wyposażone w dwa otwory: owalny (dla.owalny) do przejścia trzeciej gałęzi nerwu trójdzielnego (V3) i okrągły (dla.rotundum) do przejścia drugiej gałęzi tego samego nerwu (V2). Jednak pierwszy często łączy się z przednim otworem rozdartym (w przypadku Lacerum anteris), a drugi często łączy się z klinowo-oczodołowym otworem (w przypadku Sphenorbitale), który jest jednocześnie ograniczony zarówno przez kość skrzydłowo-klinową, jak i kość oczodołową. kość klinowa leżąca przed nią. Przez tę dziurę przechodzą nerwy okoruchowe III, trochlearis IV, odwodzące VI i pierwsza gałąź nerwu trójdzielnego.
Pokrywa czaszki jest tutaj pokryta kośćmi ciemieniowymi (ciemieniowymi), które często rosną razem (u stekowców, niektórych torbaczy i kopytnych); między nimi a górną kością potyliczną układane są jeszcze dwie kości, które łączą się w jedną kość międzyciemieniową (międzyciemieniową), która w niektórych rzędach jest zachowana jako kość niezależna (walenie, torbacze, góralki, mrówki, włochate skrzydła i niektóre gryzonie), natomiast u innych rośnie razem lub z ciemieniową (u niektórych gryzoni, syren i przeżuwaczy) lub z górną kością potyliczną (u zwierząt mięsożernych, naczelnych jednokopytnych).
Przed główną kością klinową znajduje się przednia kość klinowa (praesphenoideum) z zagłębieniem dla skrzyżowania nerwów oczodołowych; po bokach, zwykle zrośniętych z nim, leżą kości oczodołowo-klinowe (orbitosphenoidea) z otworem na przejście nerwów oczodołowych (np. opticum), zwane w przypadku zespolenia z przednią i główną kość klinową, małe skrzydełka (alae parvae). Jak wspomniano powyżej, pomiędzy obydwoma skrzydłami skrzydłowymi z każdej strony znajdują się szczelinowe otwory, przez które przechodzą nerwy mięśni oka i pierwszej gałęzi nerwu trójdzielnego (od Sphenorbitale).
Na wierzchu tego rzędu kości odpowiadają kości czołowe (czołowe) pokryte powłoką - zwykle dobrze rozwinięte sparowane kości, czasami zrośnięte i niosące pręty w rogach bydlęcych, a u jeleni - konopie na poroże. Zakrywając czaszkę, kości czołowe są ograniczone od góry i od wewnątrz orbity, często tworząc procesy: przedoczodołowy (pr. Praeorbitalis i tylny-oczodołowy (pr. Postorbitalis). Czasami wokół oczu powstaje w wyniku fuzji kość jarzmowa(jugale) z kością czołową, pierścieniem okołooczodołowym (niektóre owadożerne i mięsożerne, przeżuwacze, jednokopytne, półmałpy). U małp proces oddzielania oczodołu od oczodołu przebiega tak daleko, że między oczodołem a oczodołem tworzy się ściana kostna.
W przedniej ścianie oczodołu znajduje się niewielka powłokowa kość łzowa (lacrymale), perforowana przez ujście kanału łzowego (np. Lacrymale).
W okolicy nosa w chrzęstnej torebce nosowej znajduje się kilka skostnień. Najpierw wzdłuż chrzęstnej przegrody nosowej pośrodku rozwija się przegroda kostna (lamina perpendicularis), odpowiadająca środkowej kości węchowej (mesethmoideum); po drugie, w Tylna ściana w torebce nosowej iw przedniej ścianie jamy mózgowej rozwijają się dwie boczne płytki, perforowane licznymi [tylko w dziobaku te otwory są zastąpione przez jeden (dla. olfactorium)] otworami do przejścia nerwu węchowego - płytki sitowe ( blaszka cribrosa), odpowiadająca bocznym kościom sitowym (ethmoidalia lateralia) ... Od przegrody nosowej kostnienie rozciąga się od strony grzbietowej po bokach, biegnie wzdłuż bocznych ścian torebek nosowych i kończy się na jej dnie, tworząc liczne występy po bokach wewnątrz jamy nosowej. Są to muszle węchowe (etmoturbinalia), które wraz z płytkami bocznymi i jamami o strukturze komórkowej tworzą niekiedy bardzo złożony błędnik nosowy. Położenie umywalek można zobaczyć na załączonym schematycznym rysunku (ryc. 535). Z powłokowej kości nosa (nosa), która pokrywa ten obszar od góry, w jamie nosowej zwisają, również po jednej z każdej strony, muszle (nasoturbmalia). Dół włączony przegroda nosowa niesparowana kość, która wcześniej była uważana za vomer, nakłada się na siebie. Jednak ostatnie badania wykazały, że oprócz tej niesparowanej kości powłokowej u podstawy czaszki rozwijają się dwie sparowane kości, zwane praevomerem; a ponieważ gady mają vomer w postaci sparowanej kości, a przyklinowy był kością niesparowaną w tym obszarze, to vomer ssaków można uznać za homolog tej ostatniej kości gadów, a pre-vomery są uważane za vomery. Z przodu ściany jamy nosowej to kości szczęki (szczęka), na których tworzą się muszle wrastające w jamę nosową; są to muszle szczękowe (maxilloturbinale).
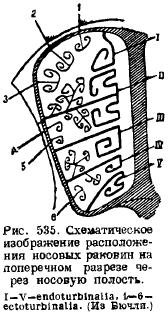
Przednia strona czaszki kończy się przed kośćmi międzyszczękowymi (praemaxillare). U małp antropomorficznych i u ludzi kość ta rośnie bardzo wcześnie wraz z kością szczęki, au nietoperzy jest zredukowana do małej kości lub w ogóle jej nie ma. Dzięki pionowemu wyrostkowi kość międzyszczękowa ogranicza przód otworu nosowego i rozciąga się między kością nosową a szczęką daleko do tyłu, czasem do kości czołowej. Od dołu kości międzyszczękowe biorą udział w tworzeniu podniebienia twardego. Ograniczają przód i boki na podniebieniu twardym otworów kanału podniebienno-posovy (stenson) (w przypadku Incisiva), łączących Jama ustna z organami Jacobsona. Wyrostki kości międzyszczękowych leżące poza tymi otworami podniebienno-nosowymi nakładają się na każdą kość niezależnie, są prawdopodobnie homologami par vomeru gadów (praevomer) i są zachowane w dziobaku jako niezależne kości. Siekacze znajdują się na kości międzyszczękowej.
Kości szczęki (szczęka), jak już wspomniano, ograniczają dno jamy nosowej, tworząc podniebienie twarde z wyrostkami poziomymi. Po wewnętrznej stronie jamy nosowej dolna powłoka nośna (maxilloturbinae) rośnie do jej bocznych ścian, utworzona przez kości szczęki, która jest ułożona jako niezależna kość.
Kontynuacją podniebienia twardego do tyłu są poziome wyrostki kości podniebiennych, ograniczające wewnętrzny kanał nosowy jako ściany boczne. Stoją pionowo, a pionowo stojące kości skrzydłowe (pterygoideum) również przylegają do nich od tyłu, które u ludzi łączą się z kością klinową, tworząc jej wyrostki skrzydłowe (proc. Pterygoidei).
Przednia część czaszki ulega znacznym zmianom w związku z przejściem do wodnego trybu życia u waleni, syren oraz w związku z rozwojem tułowia słoni.

U waleni (ryc. 536) nozdrza są cofane daleko do obszaru międzyoczodołowego i prowadzą odpowiednio do pionowego kanału, otwierającego się nozdrzami do jamy ustnej; kości nosowe leżą za nozdrzami, zachodzą na kości czołowe i prawie przylegają do kości ciemieniowej; również kości szczęk zachodzą na kości czołowe, sięgając prawie do kości ciemieniowych, które są silnie przesunięte na boki przez kość międzyciemieniową.
Syreny mają ten sam ruch nozdrzy do tyłu, ale nie tak daleko; kości nosowe są tutaj jeszcze bardziej zredukowane i mogą nawet być nieobecne (ryc. 537).

U słoni z ogromnym rozwojem kłów i potężnych zębów trzonowych oraz ze względu na obecność tułowia normalna lokalizacja kości: nozdrza są mocno cofnięte, pod tym względem kanał nosowy zajmuje prawie prostopadłe położenie; kości czołowe są bardzo krótkie, kości międzyszczękowe są bardzo duże i sięgają daleko do tyłu; kości szczęk są przesunięte pod podstawę czaszki; cała czaszka staje się bardzo masywna z niezwykle rozwiniętymi kośćmi pneumatycznymi (ryc. 538).

Dolna szczęka ssaków, w przeciwieństwie do wszystkich innych kręgowców, rozwija się z każdej strony tylko z jednej kości zębowej (zębowej); obie połówki pozostają połączone ze sobą za pomocą chrząstki, szwu lub rosną razem (zwierzęta jednokopytne, słoń, nietoperze, małpy). Każda połówka składa się z korpusu poziomego (korpusu), na którym osadzone są zęby, oraz części wznoszącej się z wyrostkami: wyrostek przedni koronalny (pros. Coronoideus), leżący za nim wyrostek stawowy (pros. Articularis) oraz u ssaków niższych, proces kątowy cofający się (ros . angularis); u większości torbaczy i niektórych gryzoni proces ten zamienia się w płytkę rozciągającą się do wewnątrz (ryc. 539).
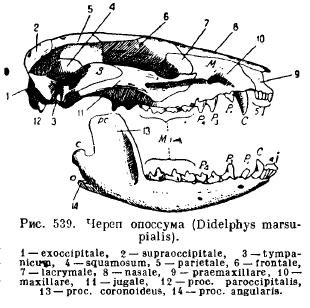
Taka budowa żuchwy i jej osobliwe przywiązanie do czaszki prowadzi nas do pytania o pochodzenie tych cech u ssaków. Historia indywidualny rozwój a dane paleontologiczne dają odpowiedź na to pytanie. W zarodkach ssaków układane są chrząstki podniebienne i chrząstki Meckeli. Jednak skostnienie chrząstki podniebienno-kwadratowej następuje poza połączeniem z przegubem żuchwy i nie prowadzi do wytworzenia kości kwadratowej, jak u gadów, a da nową kość słuchową - kowadełko, natomiast Meckela chrząstka, skostniała w tylnej części, daje kość niestawową (articulare), podobnie jak w innych szczękach, a trzecią kość słuchową stanowi młoteczek (młotek). Pierwsza kosteczka słuchowa, strzemiączko, odpowiada słupowi płazów, gadów i ptaków i, jak już wiemy, jest zmodyfikowaną zawiesiną (hyomandibulare), czyli proksymalną częścią łuku gnykowego. Podobnie jak kolumna (columella auris), strzemię zakrywa owalne okienko błędnika słuchowego i jest perforowane, jak u niektórych gadów, z otworem, przez który u niektórych niższych ssaków przechodzi tętnica twarzowa (a.facialis) (ryc. 540 i 541).

Tak więc u ssaków kość kwadratowa zamieniła się w kowadełko, kość stawowa w młoteczek; a ponieważ mała kość powłokowa rośnie do młotka, tworząc jego środkowy proces, uważa się, że ta ostatnia odpowiada kości stawowej powłoki (goniale) gadów. Leżąc obok tej ostatniej kości, powłokowa kość kątowa gadów również przechodzi w okolice ucha, stając się kością bębenkową (tympanicum). Zmiany te są widoczne na załączonym schemacie (Rys. 541).
Dane paleontologiczne potwierdzają możliwość przekształceń ukrytych powyżej. Wśród zwierząt o zębach zwierzęcych (Theriodontia) są te, u których uzębienie jest szczególnie rozwinięte, sięgając do tyłu połączenia żuchwy za pomocą kwadratowej szczoteczki. W tym samym czasie kość płaskonabłonkowa schodzi wzdłuż kości kwadratowej do tego samego stawu. Kości zębowe i płaskonabłonkowe zbliżają się więc coraz bardziej.
Wreszcie, w niektórych kościach o zębach zwierzęcych kości zębowe i płaskonabłonkowe biorą udział w artykulacji wraz z kośćmi stawowymi i kwadratowymi.
Jeśli chodzi o aparat podjęzykowy ssaków, składa się on z korpusu, zmodyfikowanej części podstawy (kopuły) łuku gnykowego, rogów przednich (cornua anteriora), utworzonej przez dolną część łuku gnykowego i rogów tylnych ( róg tylny), przekształcony z pierwszego łuku skrzelowego. Rogi przednie składają się z kilku skostnień (do czterech); segment skierowany w stronę czaszki często rośnie do kości kamienistej, tworząc wyrostek rylcowaty (proc. styloideus).
Drugi i trzeci łuk skrzelowy dał początek chrząstce tarczycy (cartilago thyreoidea).
Pasy kończynowe. Obręcz barkowa ssaków charakteryzuje się przede wszystkim redukcją kości kruczych (procoracoideum, epicoracoideum, coracoideum), tendencją do zanikania obojczyków (obojczyków) oraz dobrze rozwiniętą listwą łopatki (spina scapulae), przechodzącą w wyrostek barkowy (akromion).
Jednak u stekowców obręcz barkowa bardzo przypomina gady. Łopatka jest wyposażona w występ akromialny, do którego przylega obojczyk z przodu, a z tyłu łopatki mocowane są szerokie i krótkie kości kruków (coracoideum), idące do mostka. Przed nimi znajdują się szerokie prokorakoidy (procoracoideum) w postaci niezależnych kości. Obojczyki (obojczyki) są przyśrodkowo połączone z epiforą (episternum) (ryc. 542). U innych ssaków krakoidy układają się w postaci dwóch skostnień, ale tracą kontakt z mostkiem i dorastają do łopatki, tworząc wyrostek kruczy (proc. Coracoideus); prorakoidy znikają całkowicie (ryc. 543), obojczyki zanikają u niektórych drapieżników i kopytnych w wyniku przystosowania do szybkiego biegania, au waleni i syren w wyniku przekształcenia kończyn przednich w płetwy.

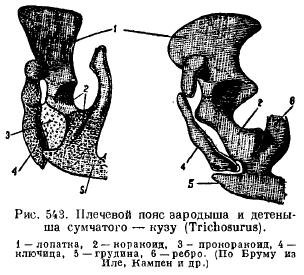
Obręcz miedniczna ssaków ma typowy układ elementów podczas układania; jego cechą jest, po pierwsze, obecność dodatkowej kości w panewce (panewce), a po drugie, zespolenie wszystkich tych kości z każdej strony w jedną bezimienną kość (innominatum) oraz zespolenie kości łonowej i kulszowej, tak że pomiędzy tymi kośćmi powstaje zamknięty staw ze wszystkich stron otworu (na przykład obturatum) (ryc. 544). U stekowców i torbaczy do kości łonowej od dołu przymocowana jest specjalna kość, która podtrzymuje worek (torsupiale).
Szkielet kończyn w pierwotnych formach ssaków wynika to z ich przystosowania do szybkich ruchów - biegania za pomocą czterech kończyn, które znajdują się pod ciałem, dzięki czemu ta ostatnia nie wisi między kończynami, jak u większości gadów, ale odpoczywa bezpośrednio na nich. Zdolność do biegania wynika z odmiennego ułożenia kończyn u ssaków. To z nimi kończyny są zorientowane tak, że stawy łokciowe kończyn przednich są skierowane do tyłu, a stawy kolanowe tylnych skierowane są do przodu. Podczas ruchu przednie kończyny są wyrzucane do przodu i ciągną ciało poprzez zginanie nadgarstka i stawy łokciowe; tylne kończyny popychają tułów do przodu, prostując nogę w stawie kolanowym.
Pod względem liczby kości tworzących kończyny i ich umiejscowienia, szkielet kończyn pradawnych form ssaków ma bardzo typową budowę prymitywnej pięciopalcowej kończyny i uległ duże zmiany w klasie dopiero w procesie adaptacji do różnego rodzaju ruchów w różne warunkiŚroda.
Kończyna przednia charakteryzuje się obecnością w dystalnej kość ramienna dziury (dla.entepicondylare), obecność proksymalnego występu kości łokciowej (olecranon), obecność kości grochowatej (pisiforme) i zespolenie czwartej i piątej dalszej kości nadgarstka (carpalia distalia 4 + 6 = hamatum, lub uncinatum) i środkowy proksymalny z pośrednim (centra dist. + intermedium = lunatum lub semilunare). Ze względu na to, że często stosuje się nomenklaturę kości nadgarstków ssaków, przyjętą w anatomii człowieka, podajemy tabelę tych nazw na schemacie wskazującym położenie kości.
Kończyna tylna ssaków charakteryzuje się przede wszystkim tym, że udo jest skręcone przez staw kolanowy przed udem i ma dobrze rozwinięte wyrostki na części proksymalnej - krętarza w liczbie 2-3 (krętarz większy, mniejszy i tertius). ); w podudzie kość piszczelowa jest bardziej rozwinięta, znajduje się kostna rzepka (rzepka), która chroni staw kolanowy. W stępie znajdują się tylko dwie bliższe kości, dobrze rozwinięte: wewnętrzna - kość skokowa (skokowa = astragalus), powstała z połączenia bliższego pośredniego i środkowego (pośrednie + bliższe środkowe) oraz kości piętowej zewnętrznej (piętowa). , odpowiadający kości strzałkowej stępu (strzałkowej). Od środkowego rzędu kości środkowa dystalna (centralna dystalna) zamieniła się w łódeczkę (naviculare = scaphoideum). Kość piszczelowa jest albo zmniejszona, albo przyczepiona do tego ostatniego.
Układ kości stępu i śródstopia oraz synonimy nazw są następujące:
W związku z przystosowaniem do ruchu w różnych środowiskach kończyny ssaków uległy bardzo istotnym zmianom, a zmiany te można szczególnie dobrze prześledzić na szkielecie. Niestety nie dysponujemy danymi paleontologicznymi dotyczącymi budowy kończyn przodków ssaków. Nie ma wątpliwości, że pierwsze ssaki miały mało wyspecjalizowanych kończyn w rodzaju opisanym powyżej. Właśnie takie kończyny najbardziej prymitywne torbacze współczesnych ssaków posiadają np. oposy – torbacze szczurów (Didelphys) oraz owadożerne prowadzące nadrzewny tryb życia. Błędem byłoby jednak wnioskować z tego, że przodkowie ssaków byli z pewnością mieszkańcami lasów, a ponadto formami wspinającymi się po drzewach. Ponadto kończyna oposa, choć służy do poruszania się po gałęziach drzew, nie jest bynajmniej typową, wyspecjalizowaną konarą drzewa, ale zachowuje swój uniwersalny pięciopalczasty typ. Takie kończyny mogły najlepiej przetrwać u zwierząt, które nie tylko biegały po ziemi, ale także wspinały się na drzewa, a także zwierzęta, które wspinały się po górach. Specjalizacja kończyn ssaków postępowała równocześnie z różnicowaniem rzędów i innych mniejszych jednostek systematycznych i nastąpiła konwergencja.
Kończyny ulegają szczególnie głębokim modyfikacjom, gdy przystosowują się do lotu, pływania w wodzie, poruszania się pod ziemią; powolne ruchy naziemne i nadrzewny tryb życia nie są związane z głęboką specjalizacją.
U nietoperzy (Chiroptera) kończyna przednia przekształca się w skrzydło, a profil rozciągnięty jest między czterema (2-b) mocno wydłużonymi palcami, pozbawionymi pazurów; pierwszy palec u nogi jest krótki i ma mocno rozwinięty pazur na końcu; ten palec nie odgrywa żadnej roli w lataniu, służy do wspinaczki; w nadgarstku kości łódeczkowate, półksiężycowate i centralne zrastają się w jedno; przedramię jest wydłużone i składa się z dobrze rozwiniętego promienia, podczas gdy kość łokciowa jest zmniejszona i narasta do promienia. Płat obejmuje również tylne nogi. Pomiędzy tymi ostatnimi różnie rozwija się u nietoperzy (ryc. 545).
U ssaków pływających w wodzie kończyny zamieniają się w płetwy w taki sam sposób, jak widzieliśmy to już u gadów. Jednocześnie szczególnie zmieniły się kończyny wielorybów i syren. W przeciwieństwie do skrzydła kończyna przednia jest znacznie skrócona w części proksymalnej. U wielorybów kości barku i przedramienia przekształciły się w krótkie, szerokie płytki, które zapewniają miejsce dla silnych mięśni. Wszystkie części kończyny są ze sobą nieruchomo połączone, tworząc mocne wiosło. W takim przypadku liczbę paliczków można znacznie zwiększyć. Te same zmiany widzieliśmy u gadów. Wzrost liczby paliczków zależy od tego, że trzon paliczków nie rośnie razem z szyszynką i kostnieje z oddzielnych ośrodków. Takie odrębne kostnienie wynika z opóźnionego procesu kostnienia u tych zwierząt. Powolność procesu kostnienia prowadzi do tego, że większość kości na ich połączeniu pozostaje chrzęstna.
Płetwy syren (Sirenia) mają zewnętrzne podobieństwo do płetw waleni: ramię i przedramię są połączone przegubowo, dzięki czemu mogą się zginać; są prawie wszystkie kości nadgarstka i wszystkie pięć palców z nieznacznie zmienioną liczbą paliczków (ryc. 546).
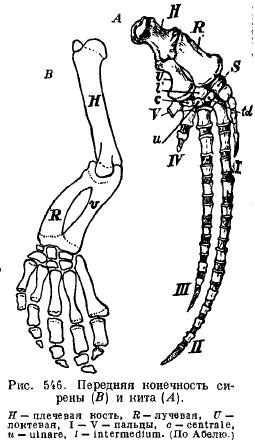
Kończyny tylne w współczesne wieloryby a syreny są zredukowane, tak że pozostają tylko resztki miednicy (ryc. 547 i 548).
Inny typ przystosowania kończyn do ruchu w palenisku obserwujemy u płetwonogich (Pinnipedia). Tutaj ogon prawie znika, a tylne nogi poruszają się do tyłu, odgrywając rolę narząd ruchu... Trzy rodziny tego rzędu reprezentują, w zależności od stopnia rozwoju zdolności przystosowania się do ruchu w wodzie, niejako trzy etapy (nie genetycznie). U morsów i fok uszatych (Otariidae) tylne kończyny mogą jeszcze wyginać się pod tułowiem do przodu, ale u morsów jest bardziej prymitywna budowa kończyn, są one krótsze i bardziej ruchliwe. U fok (Phocidae) tylne nogi nie mogą już zginać się do przodu pod tułowiem, jednak pazury na palcach są nadal zachowane (płetwa).
U zwierząt ryjących się również bardzo silnie przeobrażone są kończyny: szczególnie silnie zmieniły się kończyny torbacza (Notoryctes typhlops). Ma dwa grzbiety na łopatce, jak wiele innych zwierząt kopiących nory, co jest spowodowane silnym rozwojem mięśnia trójgłowego barku; ma obojczyk, jak wszystkie kopiące ssaki. Kość ramienna ma bardzo mocny grzbiet, który rozgałęzia się w rozwidlony kształt i służy do mocowania mocnego (niezbędnego do kopania) mięśnia naramiennego i mięśnie piersiowe; kłykieć wewnętrzny barku jest wysoko rozwinięty z powodu silny rozwój mięśnie zginaczy kości łokciowej (proc. olecranon), które służą jako punkt zaczepienia dla mięśnia trójgłowego; nadgarstek jest osobliwie zmieniony, staje się bardzo silny; Palce I i II są cofnięte w stronę dłoni, a tylko III i IV wyposażone w ogromne pazury, zwłaszcza III, służą do kopania; liczba paliczków na palcach jest bardzo mała: 2 i 1,5 śródręcza z szerokim płaskim pazurem, co zwiększa powierzchnię zagłębienia; w płaszczyźnie dłoni znajduje się kość trzeszkowa, do której przymocowany jest mięsień zginający palce, co odgrywa dużą rolę w kopaniu; zwiększa siłę nadgarstka; jak inne zakopujące się formy, spojenie kości łonowe bardzo krótki; wyrostek kulszowy rośnie wraz z wyrostkami poprzecznymi obu ostatnich kręgów krzyżowych; na udzie, kolanie i piszczeli grzbiety i wyrostki są wysoko rozwinięte - miejsca przyczepu mięśni; piszczel i strzałka (piszczel i strzałka) są mocno zrośnięte, jak złoty pieprzyk (Chrysochloris); podeszwa jest wywinięta na zewnątrz, jak w Chrysochloris; Piąty palec jest zgięty na powierzchni podeszwy, piąty paznokieć stopy jest szeroki i łopatkowaty, czwarty długi i sierpowaty, trzeci i drugi stają się krótsze i szersze; na mostku wysoko rozwinięty grzbiet, taki jak złoty pieprzyk, kretoszczur (Spalax) i kret (Talpa) (ryc. 549).
Silny rozwój wypustek na kościach w celu przyczepu mięśni, skrócenie nadgarstka, mocne połączenie nadgarstka, zmniejszenie niektórych paliczków i wzmocnienie innych, które odgrywają rolę w kopaniu - wszystkie te objawy są charakterystyczne dla ryjące ssaki w ogóle. Ale w zależności od rodzaju gleby, którą zwierzę kopie, będą różnice. Podczas kopania w miękkim podłożu (np. w kret - Talpa) (ryc. 550) ręka mocno się rozszerza, działa jak łopata i ma tendencję do rozwijania szóstego palca. Przednia kończyna kreta jest porównywalna do łopaty używanej do kopania w miękkim podłożu. Wręcz przeciwnie, w tych formach, które kopią w twardy grunt, ręka staje się węższa, tylko kilka palców bierze udział w kopaniu, najczęściej III i IV, rozwijając się silniej niż inne, jak w Notoryctes i Chrysochloris. Ta szczotka jest porównywalna do podkładki do kopania twardej ziemi.
![]()
To samo widzimy u ssaków rozrywających solidne struktury termitów, na przykład u mrówkojada (Myrmecophagus), u tamandua (Tamandua). Zwierzęta te chodzą, opierając się na zewnętrznej krawędzi łap, z wygiętymi pazurami.
Adaptacja ssaków do szybkiego poruszania się po ziemi spowodowała bardzo duże zmiany w kończynach. Ruch po ziemi, charakterystyczny dla prymitywnych ssaków, odbywał się poprzez oparcie całej dłoni i stopy; oryginalne formy ssaki były przede wszystkim plantigrade lub plantigrade, to znaczy opierały się na całych stopach podczas chodzenia, jak na przykład niedźwiedzie. Ale taka konstrukcja pozwala tylko na stosunkowo powolne chodzenie w porównaniu z szybkonogimi antylopami czy końmi, czy skaczące zające, kangury, skoczki. Podczas rozwoju filogenetycznego obserwujemy stopniowe unoszenie się ponad podłoże nadgarstka i stępu (nadgarstka i stępu), po których następowały śródręcze i śródstopie, ale paliczki nadal całkowicie dotykały podłoża. W końcu paliczki również się wyprostowały, przednie i tylne kończyny zaczęły opierać się na końcach palców. Możemy teraz krok po kroku prześledzić etapy tej ewolucji nie tylko u zwierząt kopytnych, ale także u ssaków mięsożernych, które nabyły zdolność szybkiego biegania. W miarę prostowania kończyn skrajne palce przestają dotykać podłoża, przestają funkcjonować i ulegają atrofii. Skutkuje to mniejszą powierzchnią kontaktu z podłożem. Zmiany w dystalnej części kończyn pociągały za sobą zmiany w proksymalnym: nadgarstki i stępy coraz ciaśniej przylegają do siebie, kość łokciowa i strzałkowa ulegają zmniejszeniu, kończyny zaczynają poruszać się tylko w płaszczyźnie strzałkowej, metapodia środkowe łączą się w jedna kość.
Dwie ścieżki rozwojowe prowadzą do powstania kończyn współczesnych zwierząt kopytnych. W jednym rzędzie oś kończyny przechodzi przez środek trzeciego palca i ten ostatni rozwija się najsilniej, podczas gdy reszta staje się szczątkowa. Są to koniowate (Perissodactyla), mesaksjalne (Mesaxonia). W innych oś przebiega między trzecim a czwartym palcem, rozwijając się równomiernie: parzystokopytne (Artiodactyla), przyosiowe (Paraxonia). Palce boczne są zredukowane. Rozwój koniowatych jest najwyraźniej widoczny w rozwoju filogenetycznym kończyn konia (ryc. 551). Rozwój stanu parzystokopytnego był spowodowany m.in. tym, że poprzedzony był redukcją pierwszego palca, tak że w przejściu z chodzenia pieszo prawie nie brał udziału, podczas gdy u przodków u jednego kopytnego, rozwój piątego palca był początkowo normalny.
Co ciekawe, zarówno parzystokopytne, jak i parzystokopytne wielokrotnie pojawiały się w procesie ewolucyjnym, aw niektórych przypadkach prowadziły do bardzo wyspecjalizowanego typu, na przykład u południowoamerykańskich Proterotlieriidae (wymarłych).
Pozostałe palce są pokryte kopytami, które mogą się dalej różnicować w zależności od stylu życia i siedliska. Na przykład kozy trzymające się skał mają strome i wąskie kopyta z ostrą krawędzią; w renifer są bardzo długie, szeroko rozwidlone i służą jako rodzaj nart, które zapobiegają wpadaniu zwierzęcia w śnieg i bagna.
Równolegle z palcami, wraz z rozwojem chodzenia na palcach, następuje zmiana nadgarstka (nadgarstka) i stępu (stępu). Inaczej dzieje się u parzystokopytnych i koniowatych ze względu na inny rozkład grawitacji. Wreszcie, w związku z przejściem na spacer, wartość kości łokciowej jako podparcia barku stopniowo maleje: zmniejsza się lub łączy z promieniem (promień). Podobnie kość strzałkowa jest zmniejszona i ostatecznie znika w kończynach tylnych.
Znaczenie i charakter opisanych zmian są wyraźnie widoczne z porównania kończyn wielbłąda i słonia (ryc. 552). U słonia ręka (stopa) pozostaje krótka, a ramię i przedramię (udo i podudzie) są wydłużone. Wręcz przeciwnie, u wielbłąda metapodia przyrostowe są wydłużone; ramię i przedramię (a także udo i podudzie) pozostają krótkie. Z powodu utraty wielu kości szkielet kończyny ulega uproszczeniu, ta ostatnia staje się lżejsza. Redukcja palców prowadzi do mniejszego tarcia, zmniejsza ryzyko kontuzji, zerwania więzadeł. Fuzja metapodii tworzy wielką siłę. Kończyna słonia jest znacznie cięższa. Obfitość kości w dłoni wymaga połączenia wszystkich palców w celu uzyskania stabilności. ogólna skóra... Kończyna słonia jest kolumną o wielkiej sile, ale brakuje jej lekkości, jaką ma kończyna wielbłąda.
Innym sposobem na szybkie poruszanie się po ziemi jest skakanie. Adaptacja ta pojawiała się wielokrotnie w historii ssaków, a ponadto w wielu różnych pniach filogenetycznych: u torbaczy – u kangurów (Macropus), u owadożerców – u skoczków (Macroscelidae) i wreszcie u różne stopnie rozwój gryzoni: to. zające (Leporidae) z Duplicidentata; w serii jeżozwierzy (Hystricomorplia) rozwinęła się grupa striderów (Pedetinae); wśród myszopodobnych (Myomorpha) mamy skoczek australijski (Conilurus), skoczek pustynny Starego Świata (Jaculus, Alactaga), skoczki amerykańskie (Zapus) i skoczki siatkowe (Dipodomys).
Rozwój umiejętności skakania wiąże się z wydłużeniem kończyny tylnej, czyli kości śródstopia (śródstopia). Ponadto prowadzi to do utraty dodatkowych skrajnych palców, a kości stępu są ze sobą bardzo mocno połączone. Przykładem są nogi zająca ziemnego (Alactaga) i prawdziwego skoczka (Dipus).
Przednie kończyny Alactagi są małe i krótkie. Zając ziemny używa ich podczas kopania, ale nie dotyka ziemi podczas skoku. Skakanie odbywa się tylko za pomocą tylnych nóg, a nawet bez pomocy ogona. Ten ostatni dotyka ziemi tylko w spokojnej pozycji, kiedy zwierzę spoczywa na ogonie. Szkielet nogi ma tę cechę, że kości śródstopia drugiego, trzeciego i czwartego palca łączą się w jedną kość. Końce kości śródstopia poniżej nie są zrośnięte, a środkowa (3. palec) wystaje dalej do przodu. Spośród połączonych z nimi paliczków środkowe są znowu najdłuższe. Palce I i V są szczątkowe i nie dotykają ziemi. Oba te palce mają jednakowa długość ze względu na to, że kość śródstopia kciuk dłużej niż 5. Pierwszy ma 2, drugi - 3 paliczki.
W Dipus specjalizacja poszła jeszcze dalej: pierwszy palec zniknął bez śladu, z piątego pozostał tylko szczątek. Środkowy palec dwa razy słabsze od dwóch skrajnych i tylko nieco dłuższe od nich. Palce są równoległe. Palec środkowy jest zmniejszony, ponieważ jego funkcję przejmują 2 i 4, na końcach których kładzie się nacisk podczas siedzenia i skakania. W dalszej ewolucji trzeci palec u nogi powinien zniknąć: mamy niedokończony typ adaptacji w Dipus.