Woraus besteht die Brustwirbelsäule von Säugetieren? Äußerer Aufbau von Säugetieren, Skelett und Muskulatur am Beispiel eines Haushundes
Die Wirbelsäule besteht aus den Hals-, Brust-, Lenden-, Kreuzbein- und Schwanzabschnitten. Sein charakteristisches Merkmal- Platyceliale (mit flachen Oberflächen) Form der Wirbel, zwischen denen sich Knorpel befinden Bandscheiben. Die oberen Bögen sind gut definiert. Im Halsbereich gibt es sieben Wirbel, deren Länge die Länge des Halses bestimmt; nur die Seekuh und das Faultier - Choloepus hof-fmani - haben 6 davon und das Faultier - Bradypus 8-10. Die Halswirbel sind bei Giraffen sehr lang und bei Walen, die keine Halswirbelsäule haben, sehr kurz. Die Rippen sind an den Brustwirbeln befestigt und bilden den Brustkorb. Das Brustbein, das es abschließt, ist flach und nur Fledermäuse und grabende Arten mit kräftigen Vorderbeinen (z. B. Maulwürfe) haben einen kleinen Kamm (Kiel), der als Befestigungspunkt für die Brustmuskulatur dient. Im Brustbereich gibt es 9-24 (normalerweise 12-15) Wirbel, die letzten 2-5 Brustwirbel tragen falsche Rippen, die nicht bis zum Brustbein reichen. Im Lendenbereich gibt es 2 bis 9 Wirbel; Rudimentäre Rippen verschmelzen mit ihren großen Querfortsätzen. Der Sakralabschnitt besteht aus 4–10 verschmolzenen Wirbeln, von denen nur die ersten beiden wirklich sakral sind und der Rest kaudal ist. Die Anzahl der freien Schwanzwirbel reicht von 3 (beim Gibbon) bis 49 bei der Schwanzechse.
Der Grad der Beweglichkeit einzelner Wirbel variiert. Bei kleinen Lauf- und Klettertieren ist die Wirbelsäule über die gesamte Länge groß, so dass sich ihr Körper nach innen beugen kann verschiedene Richtungen und sich sogar zu einer Kugel zusammenrollen. Bei großen, sich schnell bewegenden Tieren sind die Wirbel im Brust- und Lendenbereich weniger beweglich. Bei Säugetieren, die weiterziehen Hinterbeine ah (Kängurus, Springmäuse, Springer), die größten Wirbel befinden sich an der Schwanzbasis und am Kreuzbein, und weiter vorne nimmt ihre Größe sukzessive ab. Bei Huftieren hingegen sind die Wirbel und insbesondere ihre Dornfortsätze im vorderen Teil der Brustregion größer, wo kräftige Muskeln des Halses und teilweise der Vorderbeine an ihnen befestigt sind (
Äußere Struktur des Hundes
Der Hund war das erste Haustier. Der Mensch hat es in der Antike gezähmt. Der Hund half dem Urmenschen bei der Jagd und bewachte sein Zuhause. Heutzutage sind Arbeits-, Jagd- und Zierhunde sowie Mischlinge (Mischlingshunde) bekannt. Hunde helfen, die Grenzen unseres Mutterlandes und während der Großen zu schützen Vaterländischer Krieg suchte nach Verwundeten, half bei Aufklärung und Kommunikation. Wissenschaftler erforschen die lebenswichtigen Funktionen des Körpers von Hunden. Hund vor dem Menschen besuchter Raum.
Der Körper eines Hundes, beispielsweise eines Schäferhundes oder eines Huskys, ist schlank und muskulös. Seine Gliedmaßen befinden sich nicht wie bei Reptilien seitlich am Körper, sondern unter dem Körper, sodass der Körper des Tieres den Boden nicht berührt. Der Hund läuft und stützt sich mit starken Krallen auf seine Finger. Ein flexibler Hals ermöglicht eine größere Kopfbeweglichkeit. Das Maul des Tieres ist durch bewegliche Lippen begrenzt – oben und unten. Über Oberlippe Es gibt eine Nase mit zwei äußeren Nasenöffnungen – Nasenlöchern. Die Augen haben gut entwickelte Augenlider. Die Nickhaut (drittes Augenlid) ist beim Hund, wie bei allen Säugetieren, unterentwickelt. Von allen Tieren gibt es nur Säugetiere Ohrmuschel - Ohrmuschel. Das Außenohr eines Hundes ist groß und beweglich.
Abdeckungen von Säugetieren
Die Haut von Säugetieren ist stark und elastisch, bei den meisten Tieren befinden sich darin die Haarbasen, die ein charakteristisches Merkmal dieser Wirbeltierklasse darstellen. Haaransatz . Es gibt dicke und lange Haare — Granne und kürzere, weichere - Unterfell, oder Grundierung. Die raue und kräftige Granne schützt Unterwolle und Haut vor Beschädigungen. Die luftspeichernde Unterwolle speichert die Körperwärme gut. Neben Granne und Unterfell entwickeln sich Tiere großes Haar- Tastorgane.
Die Haare von Säugetieren bestehen wie die Federn von Vögeln und die Schuppen von Reptilien aus Hornsubstanz. Zweimal im Jahr haart ein Hund wie viele andere Säugetiere – ein Teil seiner Haare fällt aus und wird durch neue ersetzt. Die Haarbasis befindet sich in einem speziellen Haarfollikel, in den die Kanäle benachbarter Haare münden. Talgdrüse. Ihre Sekrete schmieren Haut und Haare, machen sie elastisch und verhindern, dass sie durch Wasser benetzt werden.
Abbildung: Struktur der Haut und Haare eines Säugetiers
In der Haut der meisten Tiere befinden sich und Schweißdrüsen . Der Schweiß verdunstet von der Körperoberfläche und kühlt ihn. Zusammen mit dem Schweiß werden auch überschüssiges Salz und Harnstoff aus dem Körper entfernt; Somit spielen Schweißdrüsen eine Rolle zusätzliche Organe Entladung.
Bei einem Hund Schweißdrüsen ist klein und eine Kühlung des Körpers wird durch eine verstärkte Atmung erreicht.
An den Enden der Finger von Säugetieren befinden sich Hornklauen, Nägel oder Hufe. Manchmal bilden sich auch Hornformationen am Kopf (Hörner bei Nashörnern, Antilopen, Rindern etc.) oder am Schwanz (z. B. Hornschuppen bei Ratten).
Säugetierskelett
Das Skelett von Säugetieren besteht aus den gleichen Teilen wie das Skelett anderer Wirbeltiere. Der Schädel von Tieren zeichnet sich durch einen größeren Schädel aus, der damit verbunden ist große Größen Gehirn. Es ist sehr typisch für Säugetiere, dass sie 7 Halswirbel haben. Sowohl Langhalsgiraffen als auch Wale haben die gleiche Anzahl an Halswirbeln. Die Brustwirbel (normalerweise 12-15 davon) bilden zusammen mit den Rippen und dem Brustbein eine starke Brust. Massive Wirbel Lendengegend beweglich miteinander verbunden. In diesem Abschnitt kann sich der Oberkörper beugen und beugen. Die Anzahl der Lendenwirbel ist unterschiedlich verschiedene Typen(2-9), ein Hund hat 6 davon. Die Kreuzwirbelsäule (3-4 Wirbel) verschmilzt mit den Beckenknochen. Die Anzahl der Wirbel im Schwanzbereich (von drei bis mehreren Dutzend) hängt von der Länge des Schwanzes ab.

Abbildung: Säugetierskelett am Beispiel eines Hundes

Muster: Hundeschädel
Vorderbeingürtel Säugetiere bestehen aus zwei Schulterblättern mit daran befestigten Krähenknochen und zwei Schlüsselbeinen. Die Schlüsselbeine des Hundes sind nicht entwickelt. Gürtel für die Hinterbeine- Becken - besteht aus drei Paaren Beckenknochen. Die Skelette der Gliedmaßen verschiedener Säugetier- und Reptilienarten sind ähnlich, die Details ihrer Struktur sind jedoch bei verschiedenen Arten unterschiedlich und hängen von den Lebensbedingungen des Tieres ab.
Säugetiermuskeln
Die meisten Säugetiere, darunter auch Hunde, verfügen über besonders ausgeprägte Rücken-, Gliedmaßen- und Gürtelmuskeln. Der Hund kann mit großen Sprüngen laufen, seinen Körper beugen und strecken und sich abwechselnd mit den Vorder- und Hinterbeinen vom Boden abstoßen. Starke Muskeln bewegen den Unterkiefer. Mit zusammengebissenen, mit Zähnen bewaffneten Kiefern hält der Hund seine Beute fest.

Abbildung: Hundemuskeln bewegen den Unterkiefer
Schultergürtel Säugetiere, vereinfacht gesagt, bestehen aus großen Schulterblätter, mit einem korakoiden Auswuchs und Schlüsselbein(Abb. 80, 81). Im embryonalen Entwicklungsstadium und bei erwachsenen Monotremen Korakoid ist ein eigenständiger Knochen.
Spatel befindet sich oben Brust, hat eine dreieckige Form und einen hohen Auswuchs (Grat) mit Akromial ein Prozess zum Anbringen von Muskeln. An der Stelle des Processus coracoideus und des Processus acromialis befindet sich das Schulterblatt Glenoidgrube zur Artikulation der Vorderbeine.
Schlüsselbein stabförmig dient der Verbindung von Schulter und Brustbein. Das Schlüsselbein ist bei Tieren am weitesten entwickelt, die Aufgaben mit ihren Vorderbeinen ausführen. komplexe Bewegungen(Fledermäuse, Maulwürfe, Primaten, Nagetiere usw.). Bei vielen Arten, die durch monotone Bewegungen gekennzeichnet sind (Huftiere, Hunde), fehlt das Schlüsselbein. Im Allgemeinen ist der Vorderbeingürtel über Muskeln und Bänder mit dem Achsenskelett verbunden.
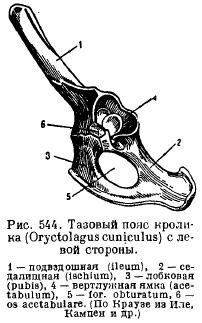
Beckengürtel (Abb. 82 A) besteht aus gepaarten namenlose Knochen, gebildet durch die Artikulation Darmbein, Sitzbein und Schambein Knochen. Becken geschlossener Typ und ist durch die Beckenknochen fest mit ihm verbunden Sakralregion Wirbelsäule. Wenn die Elemente des Beckens zusammenwachsen, kommt es dazu Schamfusion und entsteht Hüftgelenkpfanne zur Beweglichkeit der Hinterbeine. Im unteren Teil befindet sich der Innominumknochen Foramen obturatorium.
Skelett aus paarigen Gliedmaßen
Die Gliedmaßen von Säugetieren sind, wie auch die anderer Landwirbeltiere, fünffingrig und stellen einen dreigliedrigen Hebel dar, dessen Teile alle beweglich miteinander verbunden sind.
Vorderbein besteht aus Schulter, Unterarm und Hand(Abb. 81). Oberarmknochen Gut entwickelt, hat einen abgerundeten Kopf zur Befestigung am Gliedmaßengürtel. Unterarm bilden zwei Knochen - radial und ulnar, haben Olekranon oben. An der Verbindung von Schulter und Unterarm befindet sich Ellenbogengelenk.
Bürste umfasst drei Abteilungen - Handgelenk, Mittelhand und Fingerglieder. Handgelenk besteht aus 8–10 Elementen, die in drei Reihen angeordnet sind . Fessel umfasst fünf Knochen (je nach Anzahl der Finger). Jeder Finger hat drei Fingerglieder, mit Ausnahme des ersten Fingers, der aus zwei Fingergliedern besteht. Im Gegensatz zur interkarpalen Artikulation von Reptilien und Vögeln ist die bewegliche gemeinsam bei Säugetieren ist es lokalisiert zwischen Unterarm und Hand(wie Amphibien).
Hinteres Glied Auch Säugetiere haben drei Unterteilungen: Oberschenkel, Unterschenkel und Fuß(Abb. 82 B). Femur massiv und mit abgerundetem Kopf in die Hüftpfanne des Beckengürtels eingeführt. Das Schienbein enthält zwei Knochen - Schien- und Wadenbein. Das größere Schienbein bildet den inneren Teil des Unterschenkels und das Wadenbein bildet den äußeren Teil.
Zwischen Oberschenkel und Schienbein gibt es gemeinsam, bedeckt Kniescheibe, die evolutionär durch ihre verknöcherten Muskelsehnen gebildet wurde. Die Oberflächen der Gelenkknochen sind mit Knorpelformationen bedeckt. Sprunggelenk Bei Säugetieren befindet es sich zwischen den unteren Enden des Schienbeins und dem proximalen Teil des Fußes.

Reis. 82. Beckengürtel (A) und Hinterbein (B) eines Fuchses:
1 – Darmbein, 2 – Sitzbein, 3 – Schambein, 4 – Hüftpfanne, 5 – Femur, 6 – Patella, 7 – Tibia, 8 – Wadenbein, 9 – Tarsus, 10 – Mittelfuß, 11 – Phalangen, 12 – Talus, 13 – Calcaneus
Tarsalelemente die Füße sind in drei Reihen angeordnet und bilden einen Auswuchs - Kalkaneus, charakteristisch für Säugetiere. Mittelfuß besteht aus fünf Knochen, an denen die Fingerglieder befestigt sind. Wie bei den Vorderbeinen gibt es drei davon in jedem Finger, mit Ausnahme des ersten (inneren), der nur zwei Fingerglieder hat.
Säugetiere zeichnen sich durch eine besondere Anordnung der Gliedmaßen relativ zum Körper aus: Sie befinden sich unter dem Körper, was zur Entwicklung einer größeren Manövrierfähigkeit der Bewegungen beiträgt. In diesem Fall ist das Ellenbogengelenk nach hinten gerichtet und Kniegelenk, im Gegenteil, ist nach vorne ausgerichtet.
Aufgrund der Artenvielfalt der Säugetiere variieren ihre Lebens- und Bewegungsmethoden, die relative Länge der Gliedmaßen, das Verhältnis der Bestandteile, die Konfiguration der Knochen, die Anzahl der Finger usw. erheblich. Beispielsweise bei Fledermäusen Die Länge der Vorderbeine, insbesondere der Fingerglieder, ist stark verlängert und dient der Befestigung der Flugmembran.
Im Gegenteil, das Vorderbein des Maulwurfs ist verkürzt, der Handflächenteil verbreitert und der sechste Finger ist gut entwickelt. Bei Walen ist die Hand aufgrund einer Zunahme der Anzahl der Fingerglieder stark verlängert, wodurch die Gliedmaßen erworben wurden große Ähnlichkeit mit der Flosse von Lappenflossern. Die Zehen von Tieren mit großer Biomasse und schnellem Laufen (Unpaarhufer, Paarhufer) wurden deutlich reduziert und verändert.
Eines der Merkmale des Säugetierskeletts ist tolle Entwicklung Knorpel bei jungen Tieren und seine langfristige Erhaltung in der postembryonalen Entwicklungsphase. Dies wird durch die Merkmale des Knorpel- und Knochenwachstums im Zusammenhang mit dem Wachstum des Tieres selbst erklärt. Es ist der Knorpel, der mit seiner gesamten Masse (interstitiell) wächst, das Wachstum erfolgt schnell und die Proportionen der Teile können beibehalten werden, während der Knochen langsamer und nur von der Oberfläche her wächst, so dass die zum Vorschein kommenden Teile des Skeletts immer bestehen bleiben Bei gleichem Abstand bleiben die Proportionen der Teile während des Knochenwachstums nicht erhalten. Deshalb enthalten junge Tiere immer viel Knorpel, insbesondere an Stellen mit starkem Wachstum. Also, lange Knochen Gliedmaßen bestehen immer aus einem Mittelteil – der Diaphyse, die zuerst verknöchert, und Enden, die lange Zeit knorpelig bleiben – der Epiphyse. Die Verknöcherung der Diaphyse beginnt hauptsächlich an der Peripherie – perichondral, d. h. aufgrund von Zellen, die aus dem Bindegewebe stammen, das den Knorpel bedeckt – vom Perichondrium; dann kommt es zur inneren Verknöcherung des Knorpels – der endochondralen Verknöcherung; erst später, wenn durch Knochenabbau Knochenmarkhöhlen in den Diaphysen entstehen, beginnen die Epiphysen endochondral zu verknöchern; Zwischen der Diaphyse und den Epiphysen verbleibt jedoch noch lange Zeit Knorpel, mit dessen Hilfe der Knochen unter Beibehaltung seiner Proportionen in die Länge wächst (Abb. 530).
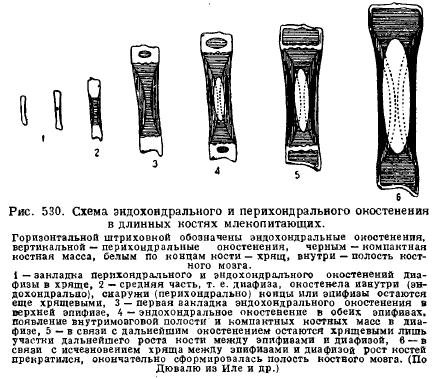
Wirbelsäule Säugetiere zeichnen sich im Gegensatz zu Sauropsida durch das Fehlen freier Gelenke zwischen den Wirbeln aus, mit Ausnahme des ersten und zweiten Wirbels. Zwischen den Wirbelkörpern liegen elastische Bindegewebsscheiben. Im Allgemeinen führt dies zu einer geringeren Flexibilität der Wirbelsäule, was auf die spezialisiertere Bewegung von Säugetieren mithilfe von Gliedmaßen zurückzuführen ist.
Die Verknöcherung der Wirbelkörper erfolgt durch die knorpeligen Epiphysen. Die Körper haben entweder flache oder leicht konvexe Oberflächen, und bei den meisten modernen Huftieren sind sie leicht opisthocoel. Die Wirbel haben gut entwickelte Dornfortsätze, insbesondere im Brustbereich, sowie Querfortsätze, die auf den Bögen sitzen; An der Basis des letzteren befinden sich Gelenkfortsätze.

Die Anzahl der Halswirbel beträgt in der Regel 7 (Abb. 531), unabhängig von der Halslänge, so dass die Halslänge im Gegensatz zu Vögeln allein von der Länge der Wirbel abhängt. Nur die Seekuh (Manatus) und eine Faultierart (Choloepus hoffmani) haben 6 Halswirbel, während Dreifingerfaultiere (Bradypus) 9 haben. Aufgrund der aquatischen Lebensweise sind sie fast ausgestorben, der Hals von Walen enthält immer noch 7 flache, teilweise verwachsene Wirbel, während im Hals einer Giraffe dieselben 7 Wirbel sehr lang sind. Die ersten beiden Halswirbel sind in Atlas und Epistropheus umgewandelt und durch echte Gelenke verbunden. Bei Monotremen fungiert der Zahnfortsatz immer noch als besonderer Knochen und ist nicht wie bei anderen Säugetieren mit dem Körper des Epistropheus verschmolzen. Die rudimentären Rippen der Halsregion verschmelzen mit den Wirbeln, wodurch perforierte Querfortsätze entstehen. Bei Walen kommt es zu einer teilweisen oder vollständigen Fusion der Halswirbel; Bei Springmäusen verschmelzen alle Halswirbel mit Ausnahme des 7., beim Beuteltier-Maulwurf (Notoryctes) verschmelzen der 2. bis 6. Wirbel, beim Maulwurf - vom 3. bis zum 4.
Die Anzahl der Brustwirbel, die durch die Befestigung von Rippen an ihnen gekennzeichnet sind, variiert von 9 [bei einem Wal (Hyperoodon) und Gürteltier (Tatusia)] bis 25 [bei einem Faultier], normalerweise sind es jedoch 12-13 . Die Brustwirbel haben gut entwickelte Fortsätze: Dornfortsätze (proc. spinosus), Quer- oder Diapophysen (proc. transversus), Gelenk- oder Zygapophysen (proc. articularis); Die Rippen sind mit ihren Köpfen am Körper eines oder zweier benachbarter Wirbel und mit ihren Tuberkeln an den Querfortsätzen befestigt. Sie bestehen aus zwei Teilen: einer knöchernen Rippe – proximal und einer knorpeligen Rippe – distal. Nur die Rippen der vorderen Brustwirbel reichen bis zum Brustbein, die hinteren Rippen enden frei (falsche Rippen). Lendenwirbel, die durch das Fehlen der Rippen gekennzeichnet sind, die hier in den Querfortsätzen enthalten sind, reichen bei Säugetieren von 2 bis 9, normalerweise sind es 6 bis 7.
Die Sakralwirbel von Säugetieren sind die Wirbel, die miteinander verwachsen artikulieren Darmbein. Es gibt 1-10 solcher Wirbel, aber nur 1-2 davon sind echte Sakralwirbel, das heißt solche, bei denen die Artikulation mit dem Becken durch unabhängige Verknöcherungen erfolgt, die den Sakralrippen entsprechen; Die übrigen Sakralwirbel sind falsche, d. h. mit dem Kreuzbein verwachsene Schwanzwirbel.
Die Anzahl der im Schwanzbereich enthaltenen Wirbel variiert enorm: von 3 ( die Fledermäuse, Gibbon) bis 49 [Eidechse (Manis macrura)].
Sternum(Sternum) von Säugetieren entsteht aus der Verschmelzung der ventralen Enden der Rippen, die dann eine unpaarige Platte bilden, die aus mehreren Abschnitten besteht (Abb. 532).
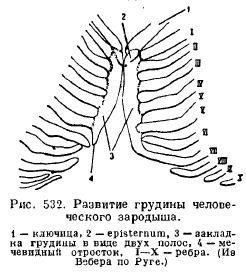
Der erweiterte vordere Teil des Brustbeins ist mit den Schlüsselbeinen (falls vorhanden) und dem ersten Rippenpaar verbunden und wird Manubrium (praesternum, manubrium) genannt; Als nächstes kommt ein Abschnitt namens Körper (Mesosternum, Corpus), der oft in eine Reihe von Knochen zerfällt, die nacheinander zwischen aufeinanderfolgenden Rippenpaaren liegen; noch weiter hinten liegt der sogenannte Schwertfortsatz (Xiphisternum). Nicht alle Rippen sind mit dem Brustbein verbunden, eine mehr oder weniger bedeutende Anzahl falscher Rippen endet frei (Abb. 533).

Bei Kloakensäugetieren (Monotremata) liegt aufgrund des Vorhandenseins von Korakoiden, die bei allen anderen Säugetieren fehlen, der prästernale Knorpel (Prosternum) vor dem Manubrium des Brustbeins, das mit den dann vorhandenen Korakoiden in Verbindung steht durch den sich entwickelnden Knochen ersetzt - das Episternum, das Kloakensäugetiere den Reptilien näher bringt.
Schädel Säugetiere weisen eine Reihe von Merkmalen auf, die sich von den Schädeln von Reptilien unterscheiden. Allerdings traten diese Merkmale nicht auf einmal auf, sondern durch allmähliche Veränderungen, was man bei der Untersuchung der Schädel fossiler Eidechsen (Theromorpha), insbesondere aus der Ordnung Theriodontia, erkennen kann. Der Säugetierschädel ist ein tropibasaler Schädel, aber mit einem verkürzten interorbitalen Teil, wodurch er wie ein platybasaler Schädel aussieht, mit einem großen Schädel, mit einer deutlichen Tendenz zur Verschmelzung einzelner Knochen.
Die Anordnung der Knochen im Schädel von Säugetieren lässt sich anhand des beigefügten Diagramms (Abb. 534) beurteilen.
An den seitlichen Hinterhauptsknochen, außerhalb der Gelenkhöcker, befindet sich normalerweise ein Hinterhauptfortsatz (proc. paroccipitalis), der bei Nagetieren und vielen Huftieren besonders gut entwickelt ist. Vor den Gelenkhöckern befindet sich normalerweise eine Öffnung für den Austritt des N. hypoglossus (XII) (z. B. Condyloideum anterius).
Im Knorpel der Gehörkapsel sind mehrere Verknöcherungen eingelagert, die jedoch schon sehr früh in zwei Knochen übergehen: den vorne liegenden – das Petrosum – und den außen und dahinter liegenden – den Warzenfortsatz (Mastoideum); Durch die Verschmelzung dieser beiden Knochen entsteht das Petromastoid oder einfach Felsenbein (Petromastoideum, Petrosum oder Perioticum), das das innere Gehörlabyrinth umschließt. Perioticum grenzt im hinteren Teil an die Haupt- und seitlichen Hinterhauptknochen und im vorderen Teil an die Keilbein- und Pterygosphenoidknochen; Da zwischen diesen Knochen und dem Felsenbein Gefäße und Nerven verlaufen, bleiben hier erhebliche Lücken: hinten - das hintere Foramen raged (für. Lacerum posterius) oder das Foramen jugularis (für. jugulare) für den Durchgang des Glossopharynx (IX ) Nerv, Äste des Vagusnervs (X) und für den Nervus accessorius (XI) sowie für die Vena jugularis interna und vorne - das vordere zerrissene Foramen (z. B. Lacerum anterius), durch das die A. carotis interna verläuft gelangt in den Schädel. Manchmal ist die Verbindung zwischen Felsenbein und Schädel so schwach, dass es zusammen mit dem daran befestigten Trommelfell leicht abgetrennt werden kann oder sogar bei der Mazeration des Schädels ganz herausfällt, wie es bei Walen der Fall ist.
Außerhalb der Ohrregion werden zwei Hautknochen gebildet. Einer davon, dessen Ursprung noch nicht ausreichend geklärt ist, entwickelt sich im Bereich des äußeren Gehörgangs in Form eines kleinen halbkreisförmigen Knochens. Dabei handelt es sich um das Paukenbein, das wahrscheinlich ein Homolog des Angulare ist. Unterkiefer Reptilien, die aufgrund von Veränderungen im Kiefergelenk zusammen mit anderen Knochen des Unterkiefers in die Ohrspeicheldrüse wanderten. Zunächst halbkreisförmig oder ringförmig im Bereich des Gehörgangs gelegen, wächst er häufig heran und bildet, meist zusammen mit dem Felsenbein und manchmal auch mit dem Schuppenbein, den äußeren Gehörgang (Meatus auditorius externus). Das Trommelfell ist über das Trommelfell gespannt. Auch tiefer im Inneren kann das Trommelfell an der Bildung der Mittelohrwände beteiligt sein. Oftmals schwillt das Trommelfell an der Unterseite der Gehörregion zu einer dünnwandigen Blase (Bulla ossea) an.
Der zweite Knochen dermaler Herkunft, der an der Bildung des Skeletts der Ohrregion beteiligt ist, ist der Schuppenknochen (Squamosum); Aufgrund der Befestigung des Unterkiefers an ihm erlangte es bei Säugetieren eine außergewöhnliche Entwicklung und Bedeutung. Bei Affen und Menschen sind alle diese Knochen der Ohrregion miteinander verschmolzen und bilden das komplexe Schläfenbein (Temporale); Der schuppige Knochen stellt hier nur einen Teil dar, nämlich die Schuppen (Squama temporalis) Schläfenbein, während das Trommelfell der Trommelfellteil (Pars tympanicum) ist und die Ohrspeicheldrüsenknorpelknochen selbst den Felsenbeinteil (Pars petrosa) und den Mastoidteil (Pars mastoidea) bilden. Der Grad der Entwicklung des Plattenepithelknochens und seine Beteiligung an der Bildung der Wände der Hirnschale und des Gehörgangs sind sehr unterschiedlich, aber das Plattenepithelknochen verfügt immer über eine Gelenkfläche (Cavitas glenoidea) zur Artikulation des Unterkiefers. Daher gibt es bei Säugetieren im Gegensatz zu allen anderen Wirbeltieren eine ganz besondere ursprüngliche Artikulation des Unterkiefers: nicht mit dem Quadratknochen, sondern mit dem Plattenepithel des Schädels. Die Gelenkfläche des Unterkiefers liegt auf der Basis, die sich von den Schuppen nach außen und vorne erstreckt Jochbeinfortsatz(proc. zygomaticus). Bei Monotremen wird es oberhalb der Gelenkfläche vom Schläfenkanal (can. temporalis) durchbohrt, was beim Schnabeltier besonders deutlich zum Ausdruck kommt. Hinter der Gelenkgrube befindet sich ein Fortsatz (proc. postglenoideus).
Vor der Hörregion wird die Schädelbasis vom Hauptkeilbein (basisphenoidale) gebildet Innenfläche die eine mehr oder weniger deutliche Vertiefung aufweist – die sogenannte Sella turcica (Sella turtica), in der sich die Hypophyse befindet. An den Seiten des Hauptkeilbeins liegen die unteren Teile der Vorderwand des Schädels, die Pterygosphenoidknochen (alisphenoidea), die meist mit diesem zu einem Knochen verschmolzen sind und große Flügel (alae magnae) dieses Knochens bilden. Die Pterygosphenoidknochen sind mit zwei Öffnungen ausgestattet: oval (für. Ovale) für den Durchgang des dritten Astes des Trigeminusnervs (V3) und rund (für. Rotundum) für den Durchgang des zweiten Astes desselben Nervs (V2). . Das erste verschmilzt jedoch oft mit dem vorderen Foramen ragata (z. B. Lacerum anteris) und das zweite geht oft mit dem Foramen sphenorbitale (z. B. sphenorbitale) über, das gleichzeitig sowohl vom Pterygosphenoidknochen als auch vom davor liegenden Augenhöhlen-Keilbeinknochen begrenzt wird Es. Durch diese Öffnung verlaufen der Okulomotorius (Oculomotorius III), der Trochlearis (Trochlearis IV), der Abducens (VI) und der erste Ast des Trigeminusnervs.
Die Schädeldecke ist hier von den Scheitelknochen (Parietale) bedeckt, die oft zusammenwachsen (bei Monotremen, einigen Beuteltieren und Huftieren); Zwischen ihnen und dem oberen Hinterhauptbein werden zwei weitere Knochen gelegt, die zu einem interparietalen Knochen (interparietale) verschmelzen, der in einigen Ordnungen als eigenständiger Knochen erhalten bleibt (Wale, Beuteltiere, Schliefer, Erdferkel, Wollflügel und einige Nagetiere), während bei anderen verschmilzt es entweder mit dem Scheitelbein (bei einigen Nagetieren, Sirenen und Wiederkäuern) oder mit dem oberen Hinterhauptbein (bei Fleischfressern, Einhufern, Primaten).
Vor dem Hauptkeilbein liegt das vordere Keilbein (Praesphenoideum) mit einer Vertiefung für das Chiasma der Augennerven; an den Seiten davon, meist damit verwachsen, liegen die Augenhöhlen-Keilbeinknochen (orbitosphenoidea) mit einer Öffnung für den Durchgang der Augenhöhlennerven (for. opticum), bei Verschmelzung mit den kleinen Flügeln (alae parvae) genannt vordere und Hauptkeilbeinknochen. Wie oben erwähnt, befinden sich zwischen den beiden Pterygoideusknochen auf jeder Seite schlitzartige Öffnungen, durch die die Nerven der Augenmuskeln und der erste Ast des Trigeminusnervs (für. Sphenorbitale) verlaufen.
Oben auf dieser Knochenreihe befinden sich die vorderen Stirnknochen (Frontale) – normalerweise gut entwickelte paarige Knochen, die manchmal miteinander verwachsen sind und bei Rindern Stäbe und bei Hirschen Stümpfe für das Geweih tragen. Die Stirnknochen, die den Schädel bedecken, begrenzen die Oberseite und das Innere der Augenhöhle und bilden oft Fortsätze: präorbital (proc. praeorbitalis) und postorbital (proc. postorbitalis). Manchmal entsteht es um die Augen herum durch Fusion Jochbein(jugale) mit dem Stirnbein, dem periorbitalen Ring (einige Insekten- und Fleischfresser, Wiederkäuer, Einhufer, Halbaffen). Bei Affen geht der Prozess der Trennung der Augenhöhle von der Schläfenpfanne so weit, dass sich zwischen Augenhöhle und Schläfenpfanne eine Knochenwand bildet.
In der Vorderwand der Orbita liegt ein kleines integumentäres Tränenknöchelchen (lacrymale), das durch die Öffnung des Tränenkanals (for. lacrymale) perforiert ist.
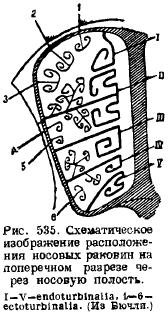
Im Nasenbereich bilden sich mehrere Verknöcherungen in der knorpeligen Nasenkapsel. Erstens entwickelt sich entlang der knorpeligen Nasenscheidewand in der Mitte eine Knochenscheidewand (Lamina Verticalis), die dem mittleren Riechbein (Mesethmoideum) entspricht; zweitens, in Rückwand Nasenkapsel und in der Vorderwand der Gehirnhöhle entwickeln sich zwei seitliche Platten, die von zahlreichen [nur beim Schnabeltier werden diese Löcher durch eine (für. Olfactorium) ersetzt] Öffnungen für den Durchgang des Riechnervs durchbrochen sind - cribriforme Platten (Lamina). cribrosa), entsprechend den seitlichen Siebbeinknochen (Ethmoidalia lateralia). Von der Nasenscheidewand aus breitet sich die Verknöcherung von dorsal an den Seiten aus, verläuft entlang der Seitenwände der Nasenkapseln und endet unten, wobei sie zahlreiche Vorsprünge von den Seiten in die Nasenhöhle bildet. Dabei handelt es sich um Riechmuscheln (Ethmoturbinalia), die zusammen mit den seitlichen Platten und Hohlräumen der Zellstruktur ein teilweise sehr komplexes Nasenlabyrinth bilden. Die Lage der Schalen ist aus der beigefügten schematischen Zeichnung (Abb. 535) ersichtlich. Von der Haut des Nasenbeins (Nasale), die diesen Bereich nach oben abdeckt, hängen die Muscheln (Nasoturbmalia) in die Nasenhöhle, ebenfalls eine auf jeder Seite. Von unten an Nasenscheidewand liegt über einem unpaarigen Knochen, der früher als Vomer galt. Neuere Studien haben jedoch gezeigt, dass sich zusätzlich zu diesem unpaarigen Hautknochen zwei paarige Knochen, der sogenannte Prävomer, an der Schädelbasis entwickeln; und da bei Reptilien der Vomer in Form eines paarigen Knochens vorliegt und der ungepaarte Knochen in diesem Bereich das Parasphenoid war, kann der Vomer von Säugetieren als Homolog dieses letzten Knochens bei Reptilien angesehen werden, und die Vorgänger können in Betracht gezogen werden Vomer. Vor den Wänden der Nasenhöhle liegen die Kieferknochen (Maxillare), an denen sich Schalen bilden, die in die Nasenhöhle hineinwachsen; das sind die Kieferschalen (maxilloturbinale).
Die Vorderseite des Schädels endet vorne mit den Prämaxillarknochen (praemaxillare). Bei anthropomorphen Affen und Menschen verschmilzt dieser Knochen sehr früh mit dem Kieferknochen, bei Chiropteren ist er auf einen kleinen Knochen reduziert oder fehlt sogar ganz. Mit seinem vertikalen Fortsatz begrenzt das Prämaxillare die Nasenöffnung nach vorne und erstreckt sich zwischen dem Nasenbein und dem Kieferknochen weit nach hinten, manchmal bis zum Stirnbein. Von unten sind die Prämaxillarknochen an der Bildung des harten Gaumens beteiligt. Sie begrenzen die vordere und seitliche Öffnung des Gaumenkanals (Stenson-Kanals) (für Incisiva) am harten Gaumen und verbinden ihn Mundhöhle mit Jacobsons Orgel. Die Fortsätze der außerhalb dieser Palatonasalöffnungen liegenden Prämaxillarknochen sind für jeden Knochen unabhängig überlagert, vermutlich Homologe des paarigen Vomer der Reptilien (Praevomer) und beim Schnabeltier als eigenständige Knochen erhalten. Die Schneidezähne liegen am Prämaxillarknochen.
Die Kieferknochen (Maxillare) begrenzen, wie bereits erwähnt, den Boden der Nasenhöhle und bilden den harten Gaumen mit horizontalen Fortsätzen. An der Innenseite der Nasenhöhle wächst die untere Nasenmuschel (Maxilloturbinae) an deren durch die Kieferknochen gebildeten Seitenwände heran und ist als eigenständiger Knochen ausgebildet.
Die Fortsetzung des harten Gaumenrückens bilden die horizontalen Fortsätze der Gaumenknochen (Palatinum), die als Seitenwände den inneren Nasengang begrenzen. Sie stehen senkrecht und sind an der Rückseite auch durch senkrecht stehende Pterygoideum-Knochen verbunden, die beim Menschen mit dem Keilbeinknochen verschmelzen und dessen Fortsätze pterygoideus (proc. pterygoidei) bilden.
Der vordere Teil des Schädels erfährt im Zusammenhang mit dem Übergang zum aquatischen Lebensstil bei Walen und Sirenen sowie im Zusammenhang mit der Entwicklung des Rüssels bei Elefanten erhebliche Veränderungen.

Bei Walen (Abb. 536) sind die Nasenlöcher weit nach hinten in den Interorbitalbereich verlegt und münden in einen vertikalen Kanal, der mit den Choanen in die Mundhöhle mündet; Die Nasenknochen liegen hinter den Nasenlöchern, überlappen die Stirnknochen und grenzen fast an den Zwischenparietalknochen an. Außerdem überlappen die Kieferknochen die Stirnknochen und reichen fast bis zu den Scheitelknochen, wobei sie durch den Zwischenscheitelknochen stark zu den Seiten verschoben werden.
Bei Sirenen findet die gleiche Bewegung der Nasenlöcher nach hinten statt, jedoch nicht so weit; die Nasenknochen sind hier noch stärker reduziert und können sogar fehlen (Abb. 537).

Bei Elefanten sind die Stoßzähne und kräftigen Backenzähne exorbitant entwickelt und es gibt einen Rüssel normaler Standort Knochen: Die Nasenlöcher sind stark zurückgedrängt, dadurch nimmt der Nasenkanal eine fast senkrechte Stellung ein; die Stirnknochen sind sehr kurz, die Prämaxillarknochen sind sehr groß und reichen weit nach hinten; Kieferknochen sind unter die Schädelbasis verschoben; Der gesamte Schädel wird sehr massiv mit extrem ausgeprägter Pneumatik der Knochen (Abb. 538).

Der Unterkiefer von Säugetieren besteht im Gegensatz zu allen anderen Wirbeltieren auf jeder Seite aus nur einem Zahnknochen (Dentale); beide Hälften bleiben entweder durch Knorpel, Naht miteinander verbunden oder wachsen zusammen (Einhufer, Elefanten, Fledermäuse, Affen). Jede Hälfte besteht aus einem horizontalen Körper (Corpus), auf dem die Zähne sitzen, und einem aufsteigenden Teil mit Fortsätzen: einem Processus coronoideus anterior (proc. coronoideus), einem dahinter liegenden Gelenkfortsatz (proc. articularis) und bei niederen Säugetieren a rückwärtiger Winkelfortsatz (proc. articularis). Angularis); Bei den meisten Beuteltieren und einigen Nagetieren verwandelt sich dieser Prozess in eine sich nach innen erstreckende Platte (Abb. 539).
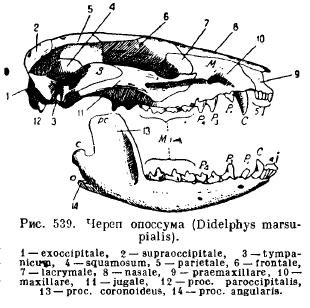
Diese Struktur des Unterkiefers und seine besondere Befestigung am Schädel führen uns zu der Frage nach dem Ursprung dieser Merkmale bei Säugetieren. Geschichte individuelle Entwicklung und paläontologische Daten geben eine Antwort auf diese Frage. In Säugetierembryonen werden das Palatoquadrat und der Meckel-Knorpel gebildet. Die Verknöcherung des Palatoquadratumknorpels erfolgt jedoch unabhängig von der Artikulation des Unterkiefers und führt nicht wie bei Reptilien zur Bildung eines Quadratknochens, sondern zur Entstehung eines neuen Gehörknochens – des Ambosses (Incus), während Meckels Knorpel, der in seinem hinteren Teil verknöchert, führt zu einem nicht artikulären Knochen (Articulare), wie bei anderen Gnathostomen, und der dritte Hörknochen ist der Hammer (Malleus). Das erste Gehörknöchelchen, Steigbügel (Stapes), entspricht der Säule von Amphibien, Reptilien und Vögeln und ist, wie wir bereits wissen, eine modifizierte Aufhängung (hyomandibulare), also der proximale Teil des Zungenbogens. Wie die Säule (columella auris) bedeckt der Steigbügel das ovale Fenster des Gehörlabyrinths und ist, wie bei einigen Reptilien, durch eine Öffnung perforiert, durch die bei einigen niederen Säugetieren die Gesichtsarterie (a. facialis) verläuft (Abb. 540). und 541).

So verwandelte sich bei Säugetieren das Quadratbein in einen Amboss, das Gelenkbein in einen Hammer; und da ein kleiner Hautknochen zum Malleus wächst und seinen Mittelfortsatz bildet, wird angenommen, dass dieser letztere dem Hautgelenkknochen (Goniale) von Reptilien entspricht. Der angrenzend an diesen letzten Knochen liegende Hautwinkelknochen der Reptilien geht auch in die Ohrregion über und wird zum Trommelfell (Tympanicum). Diese Änderungen sind im beigefügten Diagramm (Abb. 541) sichtbar.
Paläontologische Daten bestätigen die Möglichkeit der oben genannten Transformationen. Unter den Tierzahnechsen (Theriodontia) gibt es solche, bei denen der Zahnknochen eine besonders starke Entwicklung erfährt und bis hinter die Artikulationsstelle des Unterkiefers mit dem quadratischen Manus reicht. Gleichzeitig senkt sich das Squamosum entlang des Quadratum zur gleichen Artikulation. Dadurch rücken Zahn- und Plattenepithelknochen immer näher zusammen.
Schließlich sind bei einigen Säugetieren neben den Gelenk- und Quadratknochen auch die Zahn- und Plattenbeinknochen an der Artikulation beteiligt.
Der Zungenapparat von Säugetieren besteht aus einem Körper, einem modifizierten Basalteil (Copula) des Zungenbogens, Vorderhörnern (Cornua anteriora), die vom unteren Teil des Zungenbogens gebildet werden, und Hinterhörnern (Cornua posteriora). , umgewandelt aus dem ersten Kiemenbogen. Die Vorderhörner bestehen aus mehreren Verknöcherungen (bis zu vier); Das dem Schädel zugewandte Segment wächst häufig an das Felsenbein heran und bildet einen Processus styloideus (Proc. styloideus).
Der zweite und dritte Kiemenbogen führten zur Bildung des Schildknorpels (Cartilago thyreoidea).
Gliedmaßengurte. Der Schultergürtel von Säugetieren ist vor allem durch eine Verkleinerung der Rabenknochen (Procoracoideum, Epicoracoideum, Coracoideum), eine Tendenz zum Verschwinden der Clavicula und einen gut entwickelten Kamm des Schulterblatts (Spina scapulae) gekennzeichnet, der sich bis in das Akromion fortsetzt.
Allerdings ist der Schultergürtel bei Monotremen dem von Reptilien sehr ähnlich. Das Schulterblatt ist mit einem Akromialvorsprung ausgestattet, an den vorne das Schlüsselbein anschließt, und hinten sind am Schulterblatt die breiten und kurzen Rabenknochen (Coracoideum) befestigt, die zum Brustbein gehen. Vor ihnen befinden sich breite Procoracoideum (Procoracoideum) in Form unabhängiger Knochen. Die Schlüsselbeine (Clavicula) schließen medial an das Episternum an (Abb. 542). Bei anderen Säugetieren werden Krakoide in Form von zwei Ossifikationen gebildet, verlieren jedoch die Verbindung zum Brustbein und wachsen zum Schulterblatt und bilden einen Korakoidfortsatz (proc. coracoideus); die Procoracoiden verschwinden vollständig (Abb. 543), die Schlüsselbeine verschwinden bei einigen Fleischfressern und Huftieren aufgrund der Anpassung an schnelles Laufen und bei Walen und Sirenen aufgrund der Umwandlung der Vorderbeine in Flossen.

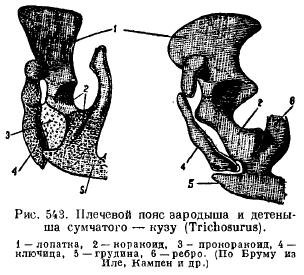
Der Beckengürtel von Säugetieren weist bei seiner Entstehung eine typische Anordnung der Elemente auf; seine Besonderheit ist erstens das Vorhandensein eines zusätzlichen Knochens in der Hüftpfanne (acetabulare), zweitens die Verschmelzung aller dieser Knochen auf jeder Seite zu einem namenlosen Knochen (innominatum) und die Verschmelzung der Schambein- und Sitzbeinknochen, so dass a Auf allen Seiten bildet sich eine geschlossene Struktur, zwischen diesen Knochen befindet sich ein Loch (z. B. obturatum) (Abb. 544). Bei Monotremen und Beuteltieren wird ein spezieller Knochen von unten am Schambein befestigt, um den Schleimbeutel (Beuteltier) zu stützen.
Gliedmaßenskelett Bei den Urformen der Säugetiere beruht es auf deren Anpassung an schnelle Bewegungen – das Laufen mit Hilfe von vier Gliedmaßen, die sich unter dem Körper befinden, so dass dieser nicht wie bei den meisten Reptilien zwischen den Gliedmaßen hängt, sondern direkt aufliegt ihnen. Die Fähigkeit zum Laufen hängt mit der unterschiedlichen Stellung der Gliedmaßen bei Säugetieren zusammen. Ihre Gliedmaßen sind so ausgerichtet, dass die Ellenbogengelenke der Vorderbeine nach hinten und die Kniegelenke der Hinterbeine nach vorne gerichtet sind. Bei der Bewegung werden die Vorderbeine nach vorne geworfen und ziehen den Körper durch Beugung des Handgelenks und Ellenbogengelenke; Die Hinterbeine schieben den Körper nach vorne, indem sie das Bein am Kniegelenk strecken.
In Bezug auf die Anzahl der in den Gliedmaßen enthaltenen Knochen und deren Lage weist das Skelett der Gliedmaßen der Urformen der Säugetiere eine sehr typische Struktur eines primitiven fünffingrigen Gliedmaßes auf und hat diese durchlaufen Große veränderungen innerhalb der Klasse erst im Prozess der Anpassung an verschiedene Arten von Bewegungen in unterschiedliche Bedingungen Umfeld.
Die Vorderbeine zeichnen sich durch das Vorhandensein im distalen Abschnitt aus Humerus Löcher (z. B. entepicondylare), das Vorhandensein eines proximalen Vorsprungs der Elle (Olecranon), das Vorhandensein eines Os pisiforme (Pisiforme) und die Verschmelzung des vierten und fünften distalen Handwurzelknochens (Carpalia dis talia 4+6 = hamatum, oder uncinatum) und das zentrale proximale mit dem intermedialen (zentraler Dist. + Intermedium = Lunatum oder Semilunare). Da häufig die in der menschlichen Anatomie übernommene Nomenklatur der Knochen des Handgelenks von Säugetieren verwendet wird, präsentieren wir eine Tabelle dieser Namen in einem Diagramm, das die Lage der Knochen angibt.
Die Hinterbeine von Säugetieren zeichnen sich vor allem dadurch aus, dass das Kniegelenk nach vorne gedreht ist, der Oberschenkel hat im proximalen Teil gut entwickelte Fortsätze - Trochanter, darunter 2-3 (Trochanter Major, Minor und Tertius); Im Unterschenkel ist das Schienbein stärker entwickelt; es gibt eine knöcherne Kniescheibe (Patella), die das Kniegelenk schützt. Der Tarsus hat nur zwei proximale Knochen, die gut entwickelt sind: den inneren Talus (Talus = Astragalus), der aus der Verschmelzung des mittleren und zentralen proximalen Knochens (Intermedium + Centrale proximale) entsteht, und den äußeren Calcaneus (Calcaneus), der dem Fibularetarsal entspricht Knochen (Fibulare). Von den Gehörknöchelchen der mittleren Reihe ging das zentrale Distal (Centrale distale) in ein Navikular (Naviculare = Scaphoideum) über. Der Schienbeinknochen war entweder reduziert oder mit diesem verschmolzen.
Die Anordnung der Fußwurzel- und Mittelfußknochen sowie die Synonymie der Namen sind wie folgt:
Im Zusammenhang mit der Anpassung an Bewegungen in verschiedenen Umgebungen haben die Gliedmaßen von Säugetieren sehr bedeutende Veränderungen erfahren, die sich besonders deutlich am Skelett verfolgen lassen. Leider liegen uns keine paläontologischen Daten zum Aufbau der Gliedmaßen der Vorfahren der Säugetiere vor. Es besteht kein Zweifel, dass die ersten Säugetiere über schlecht spezialisierte Gliedmaßen der oben beschriebenen Art verfügten. Unter unseren modernen Säugetieren haben die primitivsten Beuteltiere, zum Beispiel Opossums – Beutelratten (Didelphys) und Insektenfresser, die einen Baumlebensstil führen, genau diese Gliedmaßen. Es wäre jedoch falsch, daraus zu schließen, dass die Vorfahren der Säugetiere sicherlich Waldbewohner und darüber hinaus baumkletternde Formen waren. Darüber hinaus handelt es sich bei den Gliedmaßen des Opossums, obwohl sie für die Fortbewegung entlang von Ästen verwendet werden, keineswegs um typische, spezialisierte Baumgliedmaßen, sondern behält ihren universellen fünffingrigen Typ bei. Solche Gliedmaßen konnten am besten bei Tieren erhalten bleiben, die nicht nur auf dem Boden liefen, sondern auch auf Bäume kletterten, sowie bei Tieren, die Berge bestiegen. Die Spezialisierung der Gliedmaßen von Säugetieren erfolgte gleichzeitig mit der Differenzierung von Ordnungen und anderen kleineren systematischen Einheiten, und es kam zu Konvergenzen.
Besonders tiefgreifende Veränderungen erfahren die Gliedmaßen bei der Anpassung an den Flug, das Schwimmen im Wasser und die Bewegung unter der Erde; Langsame Bewegungen am Boden und ein baumartiger Lebensstil sind nicht auf eine tiefe Spezialisierung zurückzuführen.
Bei Fledermäusen (Chiroptera) ist das Vorderbein in einen Flügel umgewandelt und die Flugmembran ist zwischen vier (2-b) stark verlängerten Fingern ohne Krallen gespannt; Der Zeigefinger ist kurz und hat am Ende eine stark entwickelte Kralle; Dieser Finger spielt beim Fliegen keine Rolle, er wird zum Klettern verwendet; Im Handgelenk verschmelzen Kahnbein, Mondbein und Zentralknochen zu einem; Der Unterarm ist verlängert und besteht aus einem gut entwickelten Radius, während die Elle reduziert ist und am Radius anliegt. Die Flugmembran bedeckt auch die Hinterbeine. Zwischen letzteren ist es bei Fledermäusen unterschiedlich entwickelt (Abb. 545).
Bei im Wasser schwimmenden Säugetieren verwandeln sich die Gliedmaßen in ähnlicher Weise in Flossen, wie wir es bereits bei Reptilien gesehen haben. Besonders dramatisch haben sich die Gliedmaßen von Walen und Sirenen verändert. Im Gegensatz zum Flügel ist die Vorderextremität in ihrem proximalen Teil deutlich verkürzt. Bei Walen sind die Knochen der Schulter und des Unterarms zu kurzen und breiten Platten geworden, die Platz für den Ansatz kräftiger Muskeln bieten. Alle Gliedmaßen sind bewegungslos miteinander verbunden und bilden ein starkes Ruder. In diesem Fall kann die Anzahl der Phalangen stark erhöht werden. Wir haben die gleichen Veränderungen bei Reptilien gesehen. Die Zunahme der Anzahl der Phalangen hängt davon ab, dass die Diaphysen der Phalangen nicht mit den Epiphysen verschmelzen und von getrennten Zentren aus verknöchern. Diese separate Verknöcherung ist auf den langsameren Verknöcherungsprozess bei diesen Tieren zurückzuführen. Die Langsamkeit des Ossifikationsprozesses führt dazu, dass die meisten Knochen an ihren Verbindungsstellen knorpelig bleiben.
Die Flossen der Sirenen (Sirenia) ähneln im Aussehen denen der Wale: Schulter und Unterarm sind so gelenkig, dass sie sich biegen lassen; es gibt fast alle Knochen des Handgelenks und aller fünf Finger mit einer leicht veränderten Anzahl von Fingergliedern (Abb. 546).
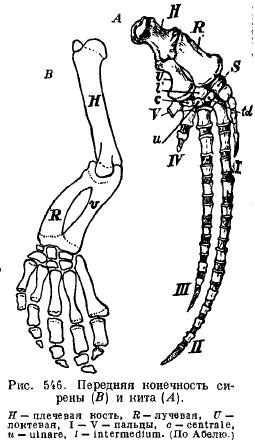
Hintere Gliedmaßen moderne Wale und Sirenen sind so stark reduziert, dass von ihnen nur noch die Reste eines Beckens übrig bleiben (Abb. 547 und 548).
Eine andere Art der Anpassung der Gliedmaßen an die Bewegung in der Schote sehen wir bei Flossenfüßern (Pinnipedia). Hier verschwindet der Schwanz fast und die Hinterbeine bewegen sich zurück und spielen die Rolle motorisches Organ. Die drei Familien dieser Ordnung repräsentieren hinsichtlich des Entwicklungsgrades der Anpassung an die Bewegung im Wasser drei Stadien (nicht genetisch). Bei Walrossen und Ohrrobben (Otariidae) können sich die Hinterbeine noch nach vorne unter dem Körper beugen, bei Walrossen ist die Struktur der Gliedmaßen jedoch primitiver, sie sind kürzer und beweglicher. Bei Robben (Phocidae) können sich die Hinterbeine nicht mehr unter dem Körper nach vorne beugen, die Zehen (Flossen) haben aber noch Krallen.
Bei grabenden Tieren unterliegen auch die Gliedmaßen sehr starken Veränderungen: Besonders stark haben sich die Gliedmaßen des Beuteltiers (Notoryctes typhlops) verändert. Es hat, wie viele grabende Tiere, zwei Leisten auf dem Schulterblatt, was auf die starke Entwicklung des Musculus triceps brachii zurückzuführen ist; Es gibt ein Schlüsselbein, wie bei allen grabenden Säugetieren. Auf dem Humerus befindet sich ein sehr starker Grat, der sich gabelförmig verzweigt und dazu dient, starke (zum Graben notwendige) Deltamuskeln zu befestigen Brustmuskeln; Der innere Kondylus des Humerus ist aufgrund dessen hoch entwickelt starke Entwicklung Ulna-Beugemuskeln (Proc. Olecranon), die als Ansatzpunkt für den Trizepsmuskel dienen; das Handgelenk ist besonders verändert und wird sehr stark; Der 1. und 2. Finger werden zur Handflächenseite bewegt, und nur der 3. und 4. Finger, ausgestattet mit riesigen Krallen, insbesondere der 3., werden zum Graben verwendet; die Anzahl der Fingerglieder ist sehr gering: 2 und 1. 5. Mittelhandknochen mit breiter, flacher Klaue, die die Grabfläche vergrößert; In der Ebene der Handfläche befindet sich ein Sesambein, an dem der Muskel befestigt ist, der die Finger beugt und beim Graben eine wichtige Rolle spielt. es erhöht die Kraft des Handgelenks; wie andere Grabformen auch die Symphyse Schambeinknochen sehr kurze; das Sitzbein verschmilzt mit den Querfortsätzen der beiden letzten Kreuzbeinwirbel; am Oberschenkel, Knie und Schienbein gibt es hoch entwickelte Grate und Auswüchse – Stellen der Muskelansätze; die Schien- und Wadenbeinknochen (Tibia und Fibula) sind fest verwachsen, wie beim Goldenen Maulwurf (Chrysochloris); die Sohle ist nach außen gedreht, wie bei Chrysochloris; Der 5. Zeh ist zur Plantarfläche gebogen, der 5. Nagel am Fuß ist breit und spatenförmig, der 4. ist lang und sichelförmig, der 3. und 2. werden kürzer und breiter; Auf dem Brustbein befindet sich ein stark entwickelter Kamm, wie der des Goldmaulwurfs, der Maulwurfsratte (Spalax) und des Maulwurfs (Talpa) (Abb. 549).
Starke Entwicklung von Leisten an den Knochen zur Muskelbefestigung, Verkürzung der Handwurzel, starke Verbindung der Handwurzeln, Reduzierung einiger Phalangen und Stärkung anderer, die beim Graben eine Rolle spielen – all diese Merkmale sind charakteristisch für grabende Säugetiere im Allgemeinen. Doch je nach Boden, in dem das Tier gräbt, werden Unterschiede zu beobachten sein. Beim Graben in weichem Boden (zum Beispiel in einem Maulwurf – Talpa) (Abb. 550) dehnt sich die Hand stark aus, sie wirkt wie eine Schaufel und neigt dazu, den 6. Finger zu entwickeln. Das Vorderbein eines Maulwurfs ist vergleichbar mit einer Schaufel, die zum Graben in weichem Boden verwendet wird. Im Gegenteil, bei den Formen, die in hartem Boden graben, wird die Hand schmaler; nur wenige Finger nehmen am Graben teil, meist der 3. und 4., die sich stärker entwickeln als andere, wie bei Notoryctes und Chrysochloris. Diese Bürste ist vergleichbar mit einem Spaten zum Graben in hartem Boden.
![]()
Dasselbe sehen wir bei Säugetieren, die die harten Strukturen von Termiten zerreißen, zum Beispiel beim Ameisenbären (Myrmecophagus) und bei den Tamandua. Beim Gehen stützen sich diese Tiere auf die äußere Kante ihrer Füße und stecken ihre Krallen ein.
Die Anpassung der Säugetiere an die schnelle Bewegung an Land hat zu sehr erheblichen Veränderungen der Gliedmaßen geführt. Die für primitive Säugetiere charakteristische Bewegung auf dem Boden erfolgte durch den Einsatz der gesamten Hand und des gesamten Fußes; Originalformen Säugetiere waren in erster Linie Plantigrade oder Plantigrade, das heißt, sie verließen sich beim Gehen auf ihren gesamten Fuß, wie zum Beispiel Bären. Eine solche Struktur ermöglicht jedoch im Vergleich zu leichtfüßigen Antilopen oder Pferden oder springenden Hasen, Kängurus und Springmäusen nur relativ langsames Gehen. Während der phylogenetischen Entwicklung sehen wir eine allmähliche Anhebung der Handwurzel und des Fußwurzelknochens (Carpus und Tarsus) über den Boden, gefolgt von der Mittelhand und dem Mittelfußknochen, aber die Phalangen berührten immer noch vollständig den Boden. Am Ende richteten sich die Fingerglieder auf, die Vorder- und Hinterbeine begannen auf den Fingerspitzen zu ruhen. Wir können die Stadien dieser Evolution nun Schritt für Schritt nicht nur bei Huftieren, sondern auch bei Raubtieren verfolgen, die die Fähigkeit zum schnellen Laufen erworben haben. Wenn sich die Gliedmaßen strecken, berühren die äußeren Zehen nicht mehr den Boden, stellen ihre Funktion ein und verkümmern. Dadurch ergibt sich eine kleinere Kontaktfläche zum Boden. Veränderungen im distalen Teil der Gliedmaßen führten zu Veränderungen im proximalen Teil: Karpalien und Tarsalien liegen immer näher beieinander, Elle und Wadenbein werden reduziert, die Gliedmaßen beginnen sich nur noch in der Sagittalebene zu bewegen, die medianen Metapodien verschmelzen in einen Knochen.
Zwei Entwicklungswege führen zur Ausbildung der Gliedmaßen moderner Huftiere. In einer Reihe verläuft die Gliedmaßenachse in der Mitte des dritten Fingers und der letzte entwickelt sich am stärksten, während der Rest rudimentär wird. Dies sind Equiden (Perissodactyla), Mesaxiale (Mesaxonia). In anderen Fällen verläuft die Achse zwischen dem dritten und vierten Finger, die sich gleichmäßig entwickeln: Artiodactyla (Artiodactyla), paraxial (Paraxonia). Die seitlichen Zehen sind reduziert. Die Entwicklung der Equiden zeigt sich am deutlichsten in der phylogenetischen Entwicklung der Gliedmaßen des Pferdes (Abb. 551). Die Entwicklung des Artiodactyl-Stadiums war unter anderem darauf zurückzuführen, dass ihm eine Reduktion der ersten Zehe vorausging, so dass diese beim Übergang vom Plantigrad-Gehen nahezu keine Rolle spielte, während sie bei den Vorfahren der Einhufer vorlag Bei Tieren war die Entwicklung der fünften Zehe zunächst normal,
Interessant ist, dass sowohl Artiodactyle als auch Equiden im Laufe der Evolution immer wieder auftraten und teilweise zu einer sehr speziellen Art führten, beispielsweise bei den südamerikanischen Proterotlieriidae (ausgestorben).
Die restlichen Zehen sind mit Hufen bedeckt, die sich je nach Lebensstil und Lebensraum weiter unterscheiden können. So haben Ziegen, die sich auf Felsen aufhalten, steile und schmale Hufe und sind mit einer scharfen Kante ausgestattet; bei Rentier Sie sind sehr lang, weit gegabelt und dienen als eine Art Ski, die verhindern, dass das Tier in den Schnee und in Sümpfe fällt.
Gleichzeitig mit den Fingern kommt es mit der Entwicklung des Gehens auf den Fingern zu einer Veränderung des Handgelenks (Carpus) und des Tarsus (Tarsus). Aufgrund einer unterschiedlichen Schwerkraftverteilung kommt es bei Artiodactylen und Equiden unterschiedlich vor. Schließlich nimmt im Zusammenhang mit dem Übergang zum Bogengehen die Bedeutung der Elle (Ulna) als Stütze für die Schulter allmählich ab: Sie nimmt ab oder geht in die Speiche (Radius) über. Auf die gleiche Weise wird das Wadenbein reduziert und verschwindet schließlich in den Hinterbeinen.
Bedeutung und Art der beschriebenen Veränderungen werden aus einem Vergleich der Gliedmaßen eines Kamels und eines Elefanten deutlich (Abb. 552). Beim Elefanten bleibt die Hand (Fuß) kurz und Schulter und Unterarm (Oberschenkel und Unterschenkel) sind verlängert. Im Gegenteil, beim Kamel verlängern sich die verschmolzenen Metapodien; Oberarm und Unterarm (sowie Ober- und Unterschenkel) bleiben kurz. Durch den Verlust einer Reihe von Knochen wird das Skelett der Extremität vereinfacht, diese wird leichter. Die Verkleinerung der Finger führt zu weniger Reibung und verringert das Risiko von Schäden und Bänderrissen. Die Verschmelzung der Metapodien schafft eine größere Stärke. Das Glied des Elefanten ist viel schwerer. Die Fülle an Knochen in der Hand erfordert die Verbindung aller Finger, um Stabilität zu gewährleisten. allgemeine haut. Das Glied eines Elefanten ist eine Säule von großer Stärke, aber es fehlt ihm die Leichtigkeit eines Kamelglieds.
Eine andere Möglichkeit, sich am Boden schnell fortzubewegen, ist das Springen. Diese Anpassung kam in der Geschichte der Säugetiere und darüber hinaus in den unterschiedlichsten phylogenetischen Stämmen immer wieder vor: bei Beuteltieren – bei Kängurus (Macropus), bei Insektenfressern – bei Springern (Macroscelidae) und schließlich bei verschiedene Reihen Entwicklung der Nagetiere: Fam. Hasen (Leporidae) aus Duplicidentata; unter den stachelschweinähnlichen Arten (Hystricomorphlia) entwickelte sich eine Gruppe langbeiniger Tiere (Pedetinae); Zu den mausähnlichen Arten (Myomorpha) zählen die Australische Springmaus (Conilurus), die Altweltspringmaus (Jaculus, Alactaga), die Amerikanische Springmaus (Zapus) und die Beutelspringmaus (Dipodomys).
Die Entwicklung der Sprungfähigkeit ist mit der Verlängerung der Hinterbeine, nämlich der Mittelfußknochen, verbunden. Darüber hinaus kommt es zum Verlust zusätzlicher Außenfinger und die Fußwurzelknochen sind sehr fest miteinander verbunden. Beispiele sind die Füße des Laufhasen (Alactaga) und der Echten Springmäuse (Dipus).
Die Vorderbeine von Alactaga sind klein und kurz. Der Erdhase nutzt sie beim Graben, berührt damit aber beim Springen nicht den Boden. Das Springen erfolgt nur mit Hilfe der Hinterbeine und auch ohne Hilfe des Schwanzes. Letzterer berührt den Boden nur in ruhiger Position, wenn das Tier auf seinem Schwanz ruht. Das Beinskelett weist die Besonderheit auf, dass die Mittelfußknochen der 2., 3. und 4. Zehe zu einem Knochen verschmelzen. Die Enden der Mittelfußknochen sind unten nicht verwachsen und der Mittelfuß (3. Zeh) ragt weiter nach vorne. Von den mit ihnen verbundenen Fingergliedern sind wiederum die mittleren die längsten. Der 1. und 5. Finger sind rudimentär und berühren den Boden nicht. Beide Finger haben die gleiche Länge aufgrund der Tatsache, dass der Mittelfußknochen Daumen länger als der 5. Das erste hat 2, das zweite 3 Phalangen.
Bei Dipus ging die Spezialisierung sogar noch weiter: Der 1. Finger verschwand spurlos, vom 5. Finger ist nur noch ein Rudiment vorhanden. Mittelfinger doppelt so schwach wie die beiden äußeren und nur geringfügig länger als diese. Die Finger sind parallel. Der Mittelfinger ist reduziert, da seine Funktion vom 2. und 4. Finger übernommen wird, deren Enden beim Sitzen und Springen Halt geben. In der weiteren Evolution muss der 3. Finger verschwinden: Bei Dipus haben wir eine unvollendete Art der Anpassung.