Šta je torakalna kičma sisara. Vanjska građa sisara, skeleta i mišića na primjeru domaćeg psa
Kičma se sastoji od vratnog, torakalnog, lumbalnog, sakralnog i kaudalnog regiona. Njegovo karakteristična karakteristika- platicelijalni (sa ravnim površinama) oblik pršljenova, između kojih se nalaze hrskavičasti intervertebralnih diskova... Gornji lukovi su dobro definisani. U cervikalnoj regiji ima sedam pršljenova čija dužina zavisi od dužine vrata; samo kod morskog krava i ljenjivca - Choloepus hof-fmani ih ima 6, a kod ljenjivca - Bradypusa 8-10. Vratni pršljenovi su vrlo dugi kod žirafa i vrlo kratki kod kitova koji nemaju vratni presjek. Rebra koja formiraju grudni koš pričvršćena su za pršljenove torakalnog regiona. Grudna kost koja ga zatvara je ravna i samo na šišmiši a vrste koje se ukopavaju sa snažnim prednjim udovima (na primjer, mladeži) imaju mali greben (kobilicu) koji služi kao tačka pričvršćivanja prsnih mišića. U torakalnom dijelu ima 9-24 (obično 12-15) pršljenova, posljednjih 2-5 torakalnih pršljenova nose - lažna rebra - ne dopiru do grudne kosti., U lumbalnoj regiji od 2 do 9 pršljenova; rudimentarna rebra se spajaju sa svojim velikim poprečnim nastavcima. Sakralni dio čine 4-10 spojenih pršljenova, od kojih su samo prva dva istinski sakralna, a ostali kaudalni. Broj slobodnih repnih pršljenova kreće se od 3 (kod gibona) do 49 kod dugorepih dinosaura.
Stepen pokretljivosti pojedinih pršljenova je različit. Kod malih životinja koje trče i penju, ona je velika, cijelom dužinom kralježnice, pa se njihovo tijelo može savijati u različitim pravcima pa čak i da se sklupčaju u klupko. Pršljenovi torakalnog i lumbalnog područja su manje pokretni kod velikih životinja koje se brzo kreću. Kod sisara koji putuju dalje zadnje noge ah (klokani, jerboasi, skakači), najveći pršljenovi nalaze se u podnožju repa i sakruma, a dalje prema naprijed njihova veličina postupno se smanjuje. Kod kopitara su, naprotiv, pršljenovi, a posebno njihovi spinovi nastavci, veći u prednjem dijelu torakalnog dijela, gdje su za njih pričvršćeni snažni mišići vrata i dijelom prednjih udova (
Vanjska struktura psa
Pas je bio prvi ljubimac. Čovjek ju je pripitomio u davna vremena. Pas je pomagao primitivnom čovjeku tokom lova, čuvao je njegov dom. Danas su poznati službeni, lovački i ukrasni psi, kao i mješanci (mješanci). Psi pomažu u obavljanju službe za zaštitu granica naše domovine, i za vrijeme Velikog Otadžbinski rat tražio ranjenike, pomagao u izviđanju i komunikaciji. Na psima naučnici provode studije vitalne aktivnosti tijela. Pas pre čoveka posjećenog prostora.
Tijelo psa, poput pastira ili haskija, je vitko i mišićavo. Njegovi udovi nisu smješteni sa strane tijela, kao kod gmizavaca, već ispod tijela, tako da tijelo životinje ne dodiruje tlo. Pas hoda oslonjen na prste sa jakim kandžama. Fleksibilan vrat omogućava odličnu pokretljivost glave. Usta životinje ograničena su pokretnim usnama - gornjom i donjom. Iznad gornja usna nalazi se nos sa par vanjskih nazalnih otvora - nozdrva. Oči imaju dobro razvijene kapke. Trepćuća membrana (treći kapak) kod psa je, kao i kod svih sisara, nedovoljno razvijena. Od svih životinja imaju samo sisari vanjskog uha - Ušna školjka... Vanjsko uho psa je veliko i pokretno.
Pokrivači sisara
Koža sisara je jaka i elastična, kod većine životinja u njoj su smještene osnove dlake koje čine karakteristiku ove klase kralježnjaka. linija kose ... Razlikovati između debelog i duga kosa — kičma i kraće, mekše - underfloor, ili poddlaka... Gruba i izdržljiva teta štiti poddlaku i kožu od oštećenja. Poddlaka, koja zadržava mnogo vazduha, dobro zadržava telesnu toplotu. Osim ostenja i donjeg krzna, razvijaju se i životinje velika kosa- organe dodira.
Dlaka sisara, poput perja ptica i krljušti reptila, sastoji se od rožnate materije. Pas se, kao i mnogi drugi sisari, dva puta godišnje linja - dio dlake mu opada i zamjenjuje se novom. Osnova kose nalazi se unutar posebne vrećice za kosu, gdje se otvaraju obližnji kanali lojne žlezde... Njihovi sekreti podmazuju kožu i kosu, što ih čini elastičnima i ne navlaženim vodom.
Uzorak: Struktura kože i dlake sisara
U koži većine životinja nalaze se i znojne žlezde ... Znoj isparava sa površine tela i hladi ga. Zajedno sa znojem, višak soli i ureje se također izlučuje iz tijela; dakle, znojne žlezde igraju ulogu dodatna tijela pražnjenje.
Pas znojne žlezde malo i hlađenje tijela postiže se pojačanim disanjem.
Na krajevima nožnih prstiju sisara nalaze se rožnate kandže, nokti ili kopita. Ponekad se rožnate formacije razvijaju i na glavi (rogovi kod nosoroga, antilopa, goveda itd.) ili na repu (na primjer, rožnate ljuske kod pacova).
Skelet sisara
Skelet sisara sastoji se od istih dijelova kao i kod drugih kralježnjaka. Lobanju životinja odlikuje se većom lobanjom, koja je povezana s velika veličina mozak. Za sisare je vrlo karakteristično prisustvo 7 vratnih pršljenova. I žirafa dugog vrata i kitovi imaju isti broj vratnih pršljenova. Torakalni pršljenovi (obično 12-15), zajedno sa rebrima i prsnom kosti, čine čvrst grudni koš. Masivni pršljenovi lumbalni pokretno artikulisani jedan s drugim. U ovom dijelu tijelo se može savijati i savijati. Broj lumbalnih pršljenova je različit u različite vrste(2-9), pas ima 6. Sakralna kičma (3-4 pršljena) se spaja sa karličnim kostima. Broj repnih pršljenova (od tri do nekoliko desetina) ovisi o dužini repa.

Slika: Kostur sisara na primjeru psa

Uzorak: pseća lobanja
Pojas prednjih nogu sisara sastoji se od dvije lopatice sa vranjskim kostima koje su za njih pričvršćene i dvije ključne kosti. Ključne kosti psa nisu razvijene. Pojas za zadnje udove- karlica - formirana od tri para karlične kosti... Skeleti udova kod različitih vrsta sisara i gmizavaca su slični, međutim, detalji njihove strukture kod različitih vrsta su različiti i ovise o životnim uvjetima životinje.
Mišići sisara
Većina sisara, uključujući psa, ima posebno razvijene mišiće leđa, udova i njihovih pojaseva. Pas može trčati velikim skokovima, savijajući i ispravljajući tijelo, naizmjenično se odričući od tla prednjim ili zadnjim nogama. Snažni mišići pokreću donju vilicu. Stezanjem čeljusti, naoružan zubima, pas čvrsto drži plijen.

Uzorak: Mišići psa koji pokreću donju vilicu
Rameni pojas sisara je pojednostavljen, sastoji se od velikog lopatice imaju korakoidni izrast, i ključna kost(sl. 80, 81). U embrionalnoj fazi razvoja i kod odraslih monotremesa coracoid je nezavisna kost.
Scapula nalazi se na vrhu prsa, trokutastog je oblika i visokog izraslina (kresta) sa akromijalni proces vezivanja mišića. Na mjestu korakoidnog izraslina i akromijalnog nastavka lopatice nalazi se glenoidna fossa za pričvršćivanje prednjeg ekstremiteta.
Ključna kost u obliku štapa služi za spajanje ramena i prsne kosti. Ključna kost je najrazvijenija kod životinja koje imaju prednje udove. složeni pokreti(šišmiši, krtice, primati, glodari, itd.). Kod mnogih vrsta, koje karakteriziraju monotoni pokreti (papkari, psi), ključna kost je odsutna. Općenito, pojas prednjih udova povezan je sa aksijalnim skeletom preko mišića i ligamenata.
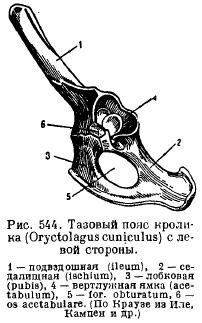
Zdjelični pojas (sl. 82 A) sastoji se od uparenih bezimene kosti artikulisan ilijačne, išijasne i stidne kosti. karlica zatvorenog tipa i kroz ilium je čvrsto povezan sa sakralni kičma. Kada elementi karlice rastu zajedno, pubic union i formirana acetabulum za pričvršćivanje stražnjih ekstremiteta. U donjem dijelu prstenaste kosti nalazi se otvor za zatvaranje.
Upareni skelet ekstremiteta
Udovi sisara, kao i kod drugih kopnenih kralježnjaka, su petoprsti i predstavljaju tročlanu polugu čiji su svi dijelovi međusobno pokretno povezani.
Prednji ud obuhvata rame, podlaktica i šaka(sl. 81). Brahijalna kost dobro razvijena, ima zaobljenu glavu za pričvršćivanje za pojas ekstremiteta. Podlakticačine dvije kosti - radijalni i ulnarni vlasništvo olecranon na vrhu. Na spoju ramena i podlaktice je lakatnog zgloba.
Četka obuhvata tri odeljenja - zglob, metakarpus i falange. Ručni zglob sastoji se od 8-10 elemenata raspoređenih u tri reda ... Pastern uključuje pet kostiju (prema broju prstiju). Svaki prst ima tri falange, sa izuzetkom prvog prsta koji ima dvije falange. Za razliku od interkarpalne artikulacije gmizavaca i ptica, mobilni joint kod sisara koji se nalaze između podlaktice i šake(kao vodozemci).
Zadnji ud sisari takođe imaju tri odeljenja - butina, potkolenica i stopalo(Sl. 82 B). Femur masivan i sa zaobljenom glavom umetnut u acetabulum karličnog pojasa. Potkolenica uključuje dve kosti - tibije i tibije. Veća tibija je unutrašnji element tibije, a fibula je njen vanjski dio.
Između butine i potkolenice nalazi se joint prikriveno kapa za koljeno koji su evoluirali iz njihovih okoštalih mišićnih tetiva. Površine zglobnih kostiju prekrivene su hrskavičnim formacijama. Zglob skočnog zgloba sisara se nalazi između donjih krajeva potkoljenice i proksimalnog dijela stopala.

Rice. 82. Karlični pojas (A) i zadnji ud (B) lisice:
1 – ilium, 2 - ischium, 3 - stidna kost, 4 - acetabulum, 5 - femur, 6 - patela, 7 - tibija, 8 - fibula, 9 - tarsus, 10 - metatarsus, 11 - falange prstiju, 12 - talus, 13 - kalkaneus
Tarzalni elementi stopala su raspoređena u tri reda i formiraju izrasline - calcaneus tipično za sisare. Metatarsus sastoji se od pet kostiju za koje su pričvršćene falange prstiju. Kao iu prednjim udovima, ima ih po tri u svakom prstu, osim prvog (unutrašnjeg), koji ima samo dvije falange.
Za sisare je karakteristično posebno pozicioniranje udova u odnosu na tijelo: oni se nalaze ispod tijela, što doprinosi razvoju veće manevarske sposobnosti pokreta. U ovom slučaju, zglob lakta je usmjeren prema stražnjoj strani, i kolenski zglob naprotiv, orijentisan je prema naprijed.
U vezi sa raznovrsnošću vrsta sisara, njihov način života i načini kretanja značajno variraju relativna dužina udova, odnos sastavnih elemenata, konfiguracija kostiju, broj prstiju itd. kod slepih miševa, dužina prednjeg uda, posebno falange prstiju, koje služe za pričvršćivanje zračne membrane.
Naprotiv, prednji ekstremitet mladeža je skraćen, palmarni dio šake je proširen, a šesti prst je dobro razvijen. Kod kitova je ruka naglo produžena zbog povećanja broja falangi prstiju, zbog čega je ud dobio velika sličnost sa perajem ribe sa poprečnim perajima. Prsti životinja s velikom biomasom i brzim trčanjem (kopinje, artiodaktili) su značajno smanjeni i modificirani.
Jedna od karakteristika skeleta sisara je veliki razvoj hrskavice kod mladih životinja i njeno dugoročno očuvanje u postembrionalnom periodu razvoja. To je zbog osobitosti rasta hrskavice i kostiju u vezi s rastom same životinje. To je hrskavica koja raste cijelom svojom masom (intersticijsko), rast se odvija brzo i proporcije dijelova se mogu sačuvati, dok kost raste sporije i samo s površine, tako da dijelovi skeleta koji se pojavljuju uvijek ostaju na istoj udaljenosti i očuvanje proporcije dijelova s rastom kosti postaje nemoguće. Zbog toga mlade životinje uvijek sadrže mnogo hrskavice, a posebno na mjestima gdje se odvija rast. dakle, duge kosti udovi se uvijek sastoje od srednjeg dijela - dijafize, koja prije svega okoštava, i krajeva, koje dugo vremena čuva hrskavična - epifiza. Osifikacija dijafize počinje prije svega s periferije - perihondralne, odnosno zbog ćelija koje potiču iz vezivnog tkiva koje prekriva hrskavicu - iz perihondrija; zatim joj se pridružuje unutrašnje okoštavanje hrskavice - endohondralno okoštavanje; tek kasnije, kada se u dijafizi pojave šupljine koštane srži raspadanjem kosti, pinealna žlijezda počinje endohondrijski okoštavati; između dijafize i epifize, međutim, dugo ostaje hrskavica, uz pomoć koje kost raste u dužinu uz očuvanje proporcija (Sl. 530).
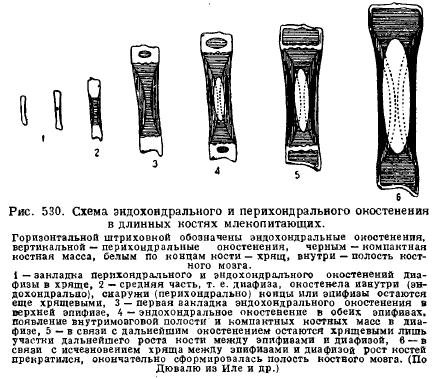
Kičma sisare karakteriše, za razliku od Sauropside, odsustvo slobodnih zglobova između pršljenova, osim prvog i drugog pršljena. Intervertebralni elastični diskovi vezivnog tkiva nalaze se između tijela pršljenova. Općenito, to rezultira manjom fleksibilnošću kralježnice, što je posljedica specijalizovanijeg kretanja sisara uz pomoć udova.
Okoštavanje tijela pršljenova događa se kroz hrskavičnu epifizu. Tijela su ili ravne površine ili blago konveksna, kod većine modernih kopitara su blago opistocela. Na kralješcima su dobro razvijeni spinozni nastavci, posebno u torakalnom dijelu, kao i poprečni nastavci koji se nalaze na lukovima; u osnovi potonjeg nalaze se zglobni procesi.

Broj vratnih pršljenova obično je 7 (sl. 531), bez obzira na dužinu vrata, pa dužina vrata, za razliku od ptica, zavisi isključivo od dužine pršljenova. Samo lamantin (Manatus) i jedna vrsta lenjivca (Choloepus hoffmani) imaju 6 vratnih pršljenova, dok troprsti lenjivci (Bradypus) imaju 9. Na vratu kitova, koji je skoro nestao zbog vodenog načina života, još uvek ima 7 ravnih pršljenova. , djelimično spojenih pršljenova, dok je u vratu žirafe ovih 7 pršljenova veoma dugačko. Prva 2 vratna pršljena su modificirana u atlas i epistrofiju, spojeni pravim zglobovima, a kod monotrema odontoidni nastavak još uvijek funkcionira u obliku posebne kosti i nije se, kao kod drugih sisara, spojio s tijelom epistrofija. Rudimentarna rebra cervikalne regije rastu zajedno s kralješcima, zbog čega se formiraju perforirani poprečni procesi. Djelomično ili potpuno spajanje vratnih pršljenova javlja se kod kitova; kod jerboa svi vratni pršljenovi rastu zajedno sa izuzetkom 7., u tobolčarskom madežu (Notoryctes) spajaju se od 2. do 6., u madežu - od 3. do 4.
Broj torakalnih pršljenova, koje karakteriziraju rebra pričvršćena za njih, varira od 9 [kod jednog kitova (Hyperoodon) i armadila (Tatusia)] do 25 [kod ljenjivca], ali obično ih ima 12-13. Procesi na torakalnim pršljenovama su dobro razvijeni: spinozni (proc. Spinosus), poprečne ili dijapofize (proc. Transversus), zglobne ili zigapofize (proc. Articularis); rebra su svojim glavama spojena s tijelom jednog ili dva susjedna pršljena i tuberkulama s poprečnim nastavcima. Sastoje se od dva dijela: koštanog rebra - proksimalnog i hrskavičnog - distalnog. Samo rebra prednjih torakalnih pršljenova dopiru do grudne kosti, zadnja rebra se završavaju slobodno (lažna rebra). Lumbalni pršljenovi, koji se odlikuju odsustvom rebara uključenih ovdje u sastav poprečnih procesa, kod sisara od 2 do 9, obično 6-7.
Sakralni pršljenovi sisara su oni pršljenovi koji su, nakon što su srasli, zglobljeni sa ilium... Takvih pršljenova ima 1-10, ali samo 1-2 od njih su pravi sakralni pršljenovi, odnosno oni kod kojih se artikulacija sa karlicom odvija kroz nezavisne osifikacije koje odgovaraju sakralnim rebrima; ostali sakralni pršljenovi su lažni, odnosno kaudalni pršljenovi pričvršćeni za sacrum.
Broj pršljenova uključenih u kaudalnu regiju značajno varira: od 3 ( šišmiši, gibon) do 49 [gušter (Manis macrura)].
Grudna kost(grudna kost) sisara nastaje iz spajanja trbušnih krajeva rebara, koji zatim formiraju jednu nesparenu ploču, koja se sastoji od više odjela (Sl. 532).
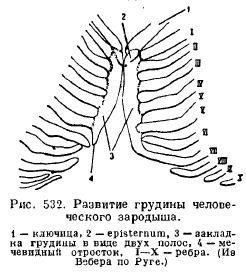
Povezan sa ključnim kostima (ako ih ima) i sa prvim parom rebara, proširena prednja grudna kost naziva se drška (praesternum, manubrium); slijedi odjel koji se zove tijelo (mesosternum, corpus), koji se često dijeli na niz kostiju, uzastopno smještenih između sljedećih parova rebara; još dalje pozadi leži takozvani xiphoid process (xiphisternum). Nisu sva rebra u vezi sa grudne kosti, manje-više značajan broj lažnih rebara se završava slobodno (Sl. 533).

Kod kloakalnih sisara (Monotremata), zbog prisustva korakoida, kojih nema kod svih ostalih sisara, ispred drške prsne kosti položena je hrskavica predgrudne kosti (prosternum), što je u vezi sa coracoids, koji se zatim pomiče kostom u razvoju - episternumom (episternmn), što kloakalnu hrskavicu približava reptilima.
Scull sisari se razlikuju po brojnim karakteristikama od lubanje gmizavaca. Međutim, ove osobine nisu se pojavile odmah, već postupnom promjenom, što se može susresti proučavanjem lubanja fosilnih zvijeri guštera (Theromorpha), posebno iz reda životinjskih zuba (Theriodontia). Lobanja sisara - tropibazalna lobanja, ali sa skraćenim interorbitalnim dijelom, zbog čega izgleda kao platibazalna, sa velikom lobanjom, sa izraženom tendencijom spajanja pojedinih kostiju zajedno,
Položaj kostiju u lobanji sisara može se suditi prema priloženom dijagramu (Sl. 534).
Na bočnim okcipitalnim kostima, prema van od zglobnih tuberkula, obično se nalazi okcipitalni izrast (pp. Paroccipitalis), koji je posebno dobro razvijen kod glodara i kod mnogih kopitara. Ispred zglobnih tuberkula obično se nalazi otvor prema vanjskoj strani hioidnog (XII) živca (for. Condyloideum anterius).
U hrskavici slušne kapsule položeno je nekoliko okoštavanja, koja se, međutim, vrlo rano spajaju u dvije kosti: ležeći sprijeda - kamenit (petrosum) i ležeći spolja i iza - mastoide (mastoideum); obje ove kosti, spojene zajedno, daju petro-mastoidnu, ili jednostavno kamenu kost (petromastoideum, petrosum ili perioticum), koja obuhvata unutrašnji slušni labirint. Perioticum se u stražnjem dijelu graniči sa glavnom i bočnom okcipitalnom kosti, au prednjem dijelu sa sfenoidnom i krilno-sfenoidnom kosti; budući da sudovi i nervi prolaze između ovih kostiju i kamene kosti, ovdje ostaju značajne praznine: iza - stražnji razderani otvor (za lacerum posterius), ili jugularni otvor (za jugulare) za prolaz glosofaringealnog (IX) živca, grane vagus (X) nerva i za pomoćni (XI) nerv, kao i za unutrašnju jugularnu venu, a sprijeda - prednji laceracijski otvor (for. Lacerum anterius), kroz koji unutrašnja karotidna arterija prolazi u lobanju . Ponekad je veza petrozne kosti sa lobanjom toliko slaba da se lako može odvojiti zajedno sa bubnjićima koja je za nju prianjala, ili čak i kada je lubanja macerirana, potpuno ispadne, kao što je slučaj kod kitova.
Izvan ušne regije formiraju se dvije integumentarne kosti. Jedan od njih, čije porijeklo još uvijek nije dovoljno razjašnjeno, razvija se u području vanjskog slušnog kanala u obliku polukružne male kosti. Ovo je bubna kost (tympanicum), koja je vjerovatno homolog kutne kosti (angulare) donja vilica gmizavci koji su se preselili u parotidnu regiju zajedno sa ostalim kostima donje vilice zbog promjena u artikulaciji vilice. Smješten u početku u obliku polukružne ili prstenaste kosti u području slušnog kanala, često raste i formira, obično zajedno sa kamenom kosti, a ponekad i s ljuskavim, vanjski slušni otvor (meatus auditorius externus). Bubna opna je rastegnuta na bubnu kost. Dublje prema unutra, bubna kost također može biti uključena u formiranje zidova srednjeg uha. Često bubna kost na donjoj strani slušne regije nabubri u mehur tankih zidova (bulla ossea).
Druga kost kožnog porijekla, koja učestvuje u formiranju skeleta ušne regije, je pločasta kost (squamosum); dobila je izuzetan razvoj i značaj kod sisara u vezi sa vezanjem donje vilice za nju. Kod majmuna i ljudi, sve ove kosti ušne regije rastu zajedno i formiraju složen temporale; pločasta kost ovdje čini samo dio, odnosno ljuske (squama temporalis) temporalna kost, dok je bubna kost bubnjić (pars tympanicum), a same parotidne hrskavične kosti čine dijelove kamenog (pars petrosa) i mastoida (pars mastoidea). Stepen razvoja skvamozne kosti i njeno učešće u formiranju zidova moždane kutije i slušnog prolaza su veoma različiti, ali pločasta kost uvek ima zglobnu površinu (cavitas glenoidea) za pričvršćivanje donje čeljusti. Dakle, sisari, za razliku od svih drugih kralježnjaka, imaju vrlo poseban originalni spoj donje čeljusti: ne za četvrtastu kost, već za skvamoznu kost lubanje. Zglobna površina donje vilice leži na bazi koja se proteže od ljuski prema van i prema naprijed zigomatski proces(proc. zygomaticus). Kod monotrema iznad zglobne površine prodire ga temporalni kanal (can.temporalis), što je posebno izraženo kod platipusa. Iza zglobne jame nalazi se proces (proc. Postglenoideus).
Ispred slušnog dela bazu lobanje čini glavna sfenoidna kost (basisphenoidale), na unutrašnja površina koja ima manje ili više značajnu depresiju - takozvano tursko sedlo (sella turtica), u kojem se nalazi hipofiza. Na stranama glavne sfenoidne kosti leže, tvoreći donje dijelove prednjeg zida lubanje, krilo-sfenoidne kosti (alisphenoidea), obično srasle s njom u jednu kost, tvoreći velika krila (alae magnae) ove kosti. Krilo-sfenoidne kosti opremljene su sa dvije rupe: ovalnom (for.ovale) za prolaz treće grane trigeminalnog živca (V3) i okruglom (for.rotundum) za prolaz druge grane istog živca (V2). Međutim, prvi se često spaja sa prednjim razderanim otvorom (for. Lacerum anteris), a drugi se često spaja sa klinasto-orbitalnim otvorom (for. Sphenorbitale), koji je istovremeno ograničen i krilo-sfenoidnom kosti i orbitalnim- sfenoidna kost koja leži ispred nje. Kroz ovu rupu prolaze nervi oculomotorius III, trochlearis IV, abducens VI i prva grana trigeminalnog živca.
Poklopac lobanje je ovdje prekriven tjemenim kostima (parietale), koje često rastu jedna s drugom (kod monotremesa, nekih torbara i kopitara); između njih i gornje okcipitalne kosti položene su još dvije kosti koje se spajaju u jednu međuparijetalnu kost (interparietale), koja je u nekim redovima očuvana kao samostalna kost (kitovi, tobolčari, hiraksi, aardvarci, vunasta krila i neki glodari), dok kod drugih raste zajedno ili s parijetalnom (kod nekih glodara, sirena i preživara), ili s gornjom potiljnom kosti (kod mesoždera, jednopapnjaka, primata).
Ispred glavne sfenoidne kosti leži prednja sfenoidna kost (praesphenoideum) sa udubljenjem za hijazmu orbitalnih nerava; na njegovim stranama, obično srasle s njim, leže orbitalno-sfenoidne kosti (orbitosphenoidea) sa otvorom za prolaz orbitalnih živaca (for. opticum), koji se naziva u slučaju srastanja s prednjom i glavnom sfenoidnom kosti, mala krila (alae parvae). Kao što je gore spomenuto, između oba pterygoida sa svake strane postoje otvori u obliku proreza kroz koje prolaze živci očnih mišića i prva grana trigeminalnog živca (for. Sphenorbitale).
Na vrhu ovog reda kostiju odgovaraju integumentarne prednje kosti (frontalne) - obično dobro razvijene parne kosti, ponekad spojene i nose štapove u goveđim rogovima, a kod jelena - konoplja za rogove. Prekrivajući lobanju, prednje kosti su ograničene odozgo i sa unutrašnje strane orbite, često formirajući procese: preorbitalni (rro. Praeorbitalis i posteriorno-orbitalni (rr. Postorbitalis). Ponekad oko očiju nastaje kao rezultat fuzije zigomatična kost(jugale) sa čeonom kosti, periobitalnim prstenom (neki insekti i mesožderi, preživari, jednopapci, polumajmuni). Kod majmuna, proces odvajanja očne duplje od temporalne duplje ide toliko daleko da se između očne duplje i temporalne duplje formira koštani zid.
U prednjem zidu orbite leži mala integumentarna suzna kost (lacrymale), perforirana otvorom suznog kanala (for. Lacrymale).
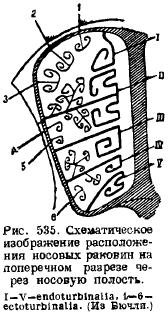
U nosnoj regiji u hrskavičnoj nosnoj kapsuli položeno je nekoliko okoštavanja. Prvo, duž hrskavičnog nosnog septuma, u sredini se razvija koštani septum (lamina perpendicularis), koji odgovara srednjoj olfaktornoj kosti (mesethmoideum); drugo, u zadnji zid u nosnoj kapsuli iu prednjem zidu moždane šupljine razvijaju se dvije lateralne ploče, perforirane brojnim [samo kod platipusa ove rupe su zamijenjene jednom (za. olfactorium)] rupama za prolaz njušnog živca - etmoidnim pločama ( lamina cribrosa), koja odgovara bočnim etmoidalnim kostima (ethmoidalia lateralia) ... Od nosnog septuma, okoštavanje se proteže od dorzalne strane duž bočnih strana, ide duž bočnih zidova nosnih kapsula i završava na njegovom dnu, formirajući brojne izbočine sa strane unutar nosne šupljine. To su mirisne školjke (ethmoturbinalia), koje zajedno s bočnim pločama i šupljinama stanične strukture ponekad tvore vrlo složen nosni labirint. Položaj sudopera može se vidjeti iz priloženog šematskog crteža (Sl. 535). Sa integumentarne nosne kosti (nasale), koja odozgo prekriva ovo područje, vise unutar nosne šupljine, takođe po jedna sa svake strane, školjke (nasoturbmalia). Na dnu nosni septum nesparena kost, koja se ranije smatrala vomerom, se preklapa. Međutim, nedavne studije su pokazale da se, pored ove nesparene integumentarne kosti, u bazi lubanje razvijaju dvije uparene kosti, nazvane praevomer; a pošto gmizavci imaju vomer u obliku uparene kosti, a parasfenoid je bio nesparena kost na ovom području, to je vomer sisara koji se može smatrati homologom ove posljednje kosti gmizavaca, a predvomeri su smatraju vomeri. Sprijeda, zidovi nosne šupljine su kosti vilice (maxillare), na kojima se formiraju školjke koje rastu u nosnu šupljinu; ovo su ljuske vilice (maxilloturbinale).
Prednja strana lubanje završava se ispred međučeljusnih kostiju (praemaxillare). Kod antropomorfnih majmuna i kod ljudi ova kost raste vrlo rano sa viličnom kosti, a kod slepih miševa je redukovana na malu kost ili čak i potpuno izostaje. Svojim okomitim procesom, intermaksilarna kost ograničava prednji dio nosnog otvora i proteže se između nosne kosti i vilice daleko unazad, ponekad do frontalne kosti. Odozdo, intermaksilarne kosti učestvuju u formiranju tvrdog nepca. Ograničavaju prednje i bočne strane na tvrdom nepcu otvora palatinsko-posovnog (stensonovog) kanala (for. Incisiva), povezujući usnoj šupljini sa Jacobsonovim orguljama. Procesi međučeljusnih kostiju koji leže izvan ovih palatinsko-nosnih otvora su superponirani za svaku kost nezavisno; oni su vjerovatno homolozi parnog vomera gmizavaca (praevomer) i očuvani su u platipusu kao samostalne kosti. Sjekutići se nalaze na intermaksilarnoj kosti.
Kosti vilice (maxillare), kao što je već spomenuto, ograničavaju dno nosne šupljine, formirajući tvrdo nepce s horizontalnim procesima. Na unutrašnjoj strani nosne šupljine na njene bočne stijenke raste donja nosiva školjka (maxilloturbinae), koju čine kosti vilice, koja je položena kao samostalna kost.
Nastavak tvrdog nepca unatrag su horizontalni nastavci palatinskih kostiju, ograničavajući unutrašnji nosni prolaz kao bočne stijenke. Stoje okomito, a sa stražnje strane im se prislanjaju i okomito stojeće pterigoidne kosti (pterygoideum), koje se kod ljudi spajaju sa sfenoidnom kosti, formirajući njene pterygoidne nastavke (proc. Pterygoidei).
Prednji dio lubanje doživljava značajne promjene u vezi s prelaskom na vodeni način života kod kitova, sirena i u vezi s razvojem trupa kod slonova.

Kod kitova (Sl. 536), nozdrve su potisnute daleko nazad u interorbitalni region i vode u vertikalni kanal, respektivno, otvarajući se hoanima u usnu šupljinu; nosne kosti leže iza nozdrva, preklapaju čeone kosti i gotovo se priklanjaju tjemenoj kosti; također kosti vilice preklapaju čeone kosti, sežu skoro do tjemenih kostiju, koje su inter-parijetalna kost snažno pomaknute u stranu.
Sirene imaju isto kretanje nozdrva unazad, ali ne tako daleko; nosne kosti su ovdje još više reducirane i mogu čak i izostati (Sl. 537).

Kod slonova s pretjerano razvijenim kljovama i snažnim kutnjacima i zbog prisustva surle, normalna lokacija kosti: nozdrve su snažno gurnute unazad, s tim u vezi, nosni kanal zauzima gotovo okomit položaj; prednje kosti su vrlo kratke, intermaksilarne kosti su vrlo velike i idu daleko unazad; kosti vilice su pomaknute ispod baze lubanje; cijela lobanja postaje vrlo masivna sa izrazito razvijenim pneumatskim kostima (Sl. 538).

Donja čeljust sisara, za razliku od svih ostalih kičmenjaka, razvija se sa svake strane samo od jedne zubne kosti (dentale); obje polovice ili ostaju povezane jedna s drugom pomoću hrskavice, šava ili rastu zajedno (jednopapke, slon, slepi miševi, majmuni). Svaka polovina se sastoji od horizontalnog tela (korpusa), na kome se nalaze zubi, i uzlaznog dela sa nastavcima: prednji krunični nastavak (rros. Coronoideus), zglobni nastavak koji leži iza njega (rros. Articularis) i kod nižih sisara, kutni proces koji ide unazad (rros . angularis); kod većine torbara i kod nekih glodara ovaj proces se pretvara u ploču koja se proteže prema unutra (Sl. 539).
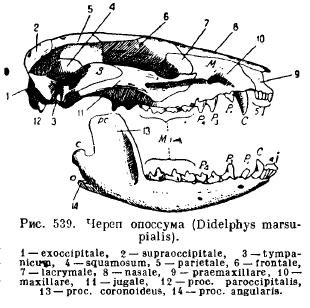
Takva struktura donje čeljusti i njeno osebujno vezivanje za lubanju dovode nas do pitanja porijekla ovih osobina kod sisara. istorija individualni razvoj a paleontološki podaci daju odgovor na to pitanje. U embrionima sisara položena je nepčana i mekelijanska hrskavica. Međutim, okoštavanje nepčane četvrtaste hrskavice nastaje izvan veze sa artikulacijom donje čeljusti i ne dovodi do stvaranja četvrtaste kosti, kao kod gmizavaca, te će dati novu slušnu kost - inkus, dok Meckelov hrskavica, okoštala u svom stražnjem dijelu, daje nezglobnu kost (articulare), kao i kod ostalih maksilarnih, a treća slušna kost je malleus (malleus). Prva slušna koščica, stapes, odgovara stubu vodozemaca, gmizavaca i ptica i, kao što već znamo, predstavlja modifikovani ovjes (hyomandibulare), odnosno proksimalni dio hioidnog luka. Poput stuba (columella auris), stremen prekriva ovalni prozor slušnog lavirinta, i perforiran je, kao kod nekih gmizavaca, sa rupom kroz koju kod nekih nižih sisara prolazi facijalna arterija (a.facialis) (Sl. 540). i 541).

Tako se kod sisara četvrtasta kost pretvorila u inkus, zglobna kost u malleus; a budući da mala integumentarna kost raste do čekića, formirajući njegov srednji nastavak, vjeruje se da ovaj posljednji odgovara integumentarnoj zglobnoj kosti (goniale) reptila. Ležeći pored ove poslednje kosti, integumentarna ugaona kost gmizavaca takođe prelazi u region uha, postajući bubna kost (tympanicum). Ove promjene su vidljive na priloženom dijagramu (Sl. 541).
Paleontološki podaci potvrđuju gore skrivenu mogućnost transformacija. Među životinjama sa zubima (Theriodontia) ima onih kod kojih je denticija posebno razvijena, koja četvrtastom četkom dopire do stražnjeg dijela spoja donje čeljusti. U isto vrijeme, pločasta kost se spušta duž četvrtaste kosti do iste artikulacije. Zubne i skvamozne kosti se, tako, sve bliže i bliže.
Konačno, kod nekih kostiju životinjskih zuba, zubne i skvamozne kosti učestvuju u artikulaciji zajedno sa zglobnim i četvrtastim kostima.
Što se tiče hipoglosalnog aparata sisara, on se sastoji od tijela, modificiranog bazalnog dijela (copula) hioidnog luka, prednjih rogova (cornua anteriora), formiranih od donjeg dijela hioidnog luka, i stražnjih rogova ( cornua posteriora), transformisana iz prvog grančičnog luka. Prednji rogovi se sastoje od nekoliko okoštavanja (do četiri); segment okrenut ka lubanji često raste do kamene kosti, formirajući stiloidni nastavak (proc. styloideus).
Drugi i treći grančasti luk doveli su do stvaranja tiroidne hrskavice (cartilago thyreoidea).
Pojasevi za ekstremitete. Rameni pojas kod sisara karakteriše prvenstveno redukcija kostiju gavrana (procoracoideum, epicoracoideum, coracoideum), tendencija nestanka ključnih kostiju (clavicula) i dobro razvijeni greben lopatice (spina scapulae), koji se nastavlja u proces akromiona. (akromion).
Međutim, kod monotreme, rameni pojas ima veliku sličnost s gmizavcima. Lopatica je opremljena akromijalnom izbočinom, uz koju je ključna kost ispred, a široke i kratke gavranove kosti (coracoideum), koje idu do grudne kosti, pričvršćene su za stražnji dio lopatice. Ispred njih se nalaze široki prokorakoidi (procoracoideum) u obliku nezavisnih kostiju. Ključne kosti (clavicula) su medijalno povezane sa epiforom (episternum) (slika 542). Kod drugih sisara, krakoidi su položeni u obliku dva okoštavanja, ali gube kontakt sa grudne kosti i rastu do lopatice, formirajući korakoidni nastavak (proc. Coracoideus); prokorakoidi potpuno nestaju (Sl. 543), ključne kosti nestaju kod nekih grabežljivaca i kopitara zbog prilagođavanja na brzo trčanje, a kod kitova i sirena zbog transformacije prednjih udova u peraje.

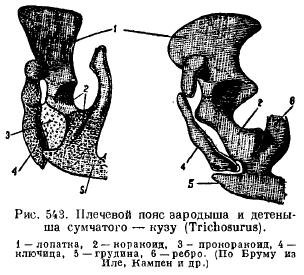
Karlični pojas kod sisara ima tipičan raspored elemenata kada je položen; njegova karakteristika je, prvo, prisustvo dodatne kosti u acetabularu (acetabulare), a drugo, fuzija svih ovih kostiju sa svake strane u jednu bezimenu kost (innominatum) i fuzija između stidne i ishijalne kosti, tako da formira se zatvoreni zglob sa svih strana rupe između ovih kostiju (for. obturatum) (sl. 544). Kod monotremesa i tobolčara, posebna kost je pričvršćena za stidnu kost odozdo za podupiranje vrećice (marsupiale).
Skelet ekstremiteta kod izvornih oblika sisara to je zbog njihove adaptacije na brze pokrete - trčanje uz pomoć četiri uda, koji se nalaze ispod tijela, tako da potonji ne visi između udova, kao kod većine gmizavaca, već miruje. direktno na njih. Sposobnost trčanja je zbog različitog položaja udova kod sisara. Kod njih su udovi orijentirani tako da su laktovi prednjih udova usmjereni prema nazad, a zglobovi koljena stražnjih usmjereni prema naprijed. Prilikom kretanja, prednji udovi se izbacuju naprijed i povlače tijelo savijanjem zgloba i lakatnih zglobova; zadnji udovi guraju trup naprijed ispravljajući nogu u zglobu koljena.
U pogledu broja kostiju koje čine udove i njihove lokacije, skelet udova predačkih oblika sisara ima vrlo tipičnu strukturu primitivnog petoprstog uda i pretrpio je velike promene unutar razreda samo u procesu prilagođavanja raznim vrstama kretanja u različitim uslovima srijeda.
Prednji ekstremitet karakterizira prisustvo u distalnom dijelu humerus rupice (for.entepicondylare), prisustvo proksimalne protruzije lakatne kosti (olecranon), prisustvo pisoformne kosti (pisiforme) i fuzija četvrte i pete distalne karpalne kosti (carpalia dis talia 4 + 6 = hamatum, ili uncinatum) i centralni proksimalni sa intermedijerom (centra dist. + intermedium = lunatum, ili semilunare). Zbog činjenice da se nomenklatura kostiju ručnog zgloba sisara, usvojena u ljudskoj anatomiji, često koristi, dajemo tablicu ovih naziva u dijagramu koji označava lokaciju kostiju.
Zadnji ekstremitet sisara karakteriše prvenstveno to što je bedro okrenuto kolenskim zglobom ispred natkolenice i ima dobro razvijene nastavke na proksimalnom dijelu - trohantera, 2-3 na broju (trohanter veliki, minor i tertius ); u potkoljenici je tibija razvijenija, nalazi se koštana patela (patela), koja štiti kolenski zglob. U tarzusu postoje samo dvije proksimalne kosti, dobro razvijene: unutrašnja - talus (talus = astragalus), nastala fuzijom srednjeg i centralnog proksimalnog (intermedium + centrale proximale), i vanjska kalkaneus (calcaneus) , što odgovara fibularnoj kosti tarzusa (fibulare). Od kostiju srednjeg reda centralna distalna (centrale distale) prešla je u skafoid (naviculare = scaphoideum). Tibijalna kost je ili smanjena ili pričvršćena za ovu potonju.
Raspored kostiju tarzusa i metatarzusa i sinonimi imena su sljedeći:
U vezi sa prilagodbom na kretanje u različitim sredinama, udovi sisara su pretrpjeli vrlo značajne promjene, a te promjene se posebno dobro mogu pratiti na skeletu. Nažalost, nemamo paleontološke podatke o građi udova predaka sisara. Nema sumnje da su prvi sisari imali male specijalizovane udove gore opisanog tipa. Upravo takve udove imaju najprimitivniji tobolčari modernih sisara, na primjer, oposumi - tobolčarski štakori (Didelphys) i insektojedi koji vode arborealni način života. Međutim, iz ovoga bi bilo pogrešno zaključiti da su preci sisara svakako bili stanovnici šuma i, osim toga, oblika koji su se penjali na drveće. Osim toga, oposumov ud, iako služi za kretanje po granama drveća, nikako nije tipičan specijalizirani ud drveta, ali zadržava svoj univerzalni tip s pet prstiju. Takvi udovi najbolje bi mogli preživjeti kod životinja koje ne samo da su trčale po zemlji, već su se pele i na drveće, kao i kod životinja koje su se penjale na planine. Specijalizacija udova sisara odvijala se istovremeno sa diferencijacijom redova i drugih manjih sistematskih jedinica, te je došlo do konvergencije.
Udovi prolaze kroz posebno duboke modifikacije kada se prilagode letu, plivanju u vodi, kretanju pod zemljom; spora kretanja tla i arborealni načini života nisu povezani s dubokom specijalizacijom.
Kod slepih miševa (Chiroptera) prednji ud je pretvoren u krilo, a aeroprofil je razvučen između četiri (2-b) jako izdužena prsta, bez kandži; prvi prst je kratak i na kraju ima snažno razvijenu kandžu; ovaj prst ne igra nikakvu ulogu u letenju, koristi se za penjanje; u zglobu, skafoidna, lunasta i centralna kost rastu zajedno u jednu; podlaktica je izdužena i sastoji se od dobro razvijenog radijusa, dok je lakatna kost smanjena i prirasla radijusu. Aeroprofil takođe zahvata zadnje noge. Između ovih, različito je razvijen kod slepih miševa (Sl. 545).
Kod sisara koji plivaju u vodi, udovi se pretvaraju u peraje na isti način kao što smo već vidjeli kod gmizavaca. Istovremeno, posebno su se promijenili udovi kitova i sirena. Za razliku od krila, prednji ud je značajno skraćen u svom proksimalnom dijelu. Kod kitova su kosti ramena i podlaktice evoluirale u kratke, široke ploče koje pružaju prostor za pričvršćivanje moćne muskulature. Svi dijelovi uda su nepomično povezani jedan s drugim, formirajući snažno veslo. U ovom slučaju, broj falangi se može znatno povećati. Videli smo iste promene kod gmizavaca. Povećanje broja falangi ovisi o činjenici da dijafiza falanga ne raste zajedno s pinealnim žlijezdama i okoštavaju iz odvojenih centara. Takvo odvojeno okoštavanje je posljedica odgođenog procesa okoštavanja kod ovih životinja. Sporost procesa okoštavanja dovodi do činjenice da većina kostiju na njihovom spoju ostaje hrskavična.
Peraje sirena (Sirenia) imaju vanjsku sličnost sa perajama kitova: rame i podlaktica su spojeni jedno s drugim, tako da se mogu savijati; ima skoro sve kosti ručnog zgloba i svih pet prstiju sa malo izmenjenim brojem falangi (sl. 546).
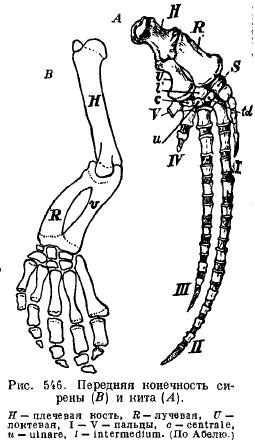
Zadnji udovi unutra moderni kitovi a sirene su smanjene tako da od njih ostaju samo ostaci karlice (sl. 547 i 548).
Još jednu vrstu prilagođavanja udova na kretanje u ognjištu vidimo kod peronožaca (Pinnipedia). Ovdje rep gotovo nestaje, a zadnje noge se pomiču unatrag, igrajući ulogu motorni organ... Tri porodice ovog reda predstavljaju, prema stepenu razvijenosti prilagodljivosti kretanju u vodi, takoreći tri stadijuma (ne genetski). Kod morževa i uhastih tuljana (Otariidae) stražnji udovi se još uvijek mogu savijati prema naprijed ispod tijela, ali kod morževa je primitivnije u građi udova, kraći su i pokretljiviji. Kod tuljana (Phocidae), zadnje noge se više ne mogu savijati prema naprijed ispod tijela, međutim, kandže su i dalje očuvane na prstima (peraja).
Kod životinja koje se ukopaju, udovi također prolaze kroz vrlo jake modifikacije: udovi tobolčarske krtice (Notoryctes typhlops) su se posebno jako promijenili. Ima dva grebena na lopatici, kao i mnoge životinje koje se ukopavaju, što je zbog snažnog razvoja mišića tricepsa ramena; ima ključnu kost, kao i svi sisari koji se kopaju. Humerus ima veoma jak greben koji se račvasto grana i služi za pričvršćivanje jakih (neophodnih za kopanje) deltoida i prsnih mišića; unutrašnji kondil ramena je visoko razvijen zbog snažan razvoj mišiće fleksora lakatne kosti (proc. olecranon), koji služi kao veza za mišić tricepsa; zglob je posebno izmijenjen, postaje vrlo jak; 1. i 2. prst su gurnuti nazad na dlan, a samo 3. i 4., opremljeni ogromnim kandžama, posebno 3., služe za kopanje; broj falangi na prstima je vrlo mali: 2 i 1. 5. metakarpal sa širokom pljosnatom kandžom, koja povećava površinu udubljenja; u ravnini dlana nalazi se sesamoidna kost, za koju je pričvršćen mišić koji savija prste, koji igra veliku ulogu u kopanju; povećava snagu zgloba; kao i drugi oblici ukopavanja, simfiza stidne kosti veoma kratko; ischium raste zajedno s poprečnim nastavcima oba posljednja sakralna pršljena; na butini, koljenu i tibiji jako su razvijeni grebeni i izrasline - mjesta pričvršćivanja mišića; tibija i fibula (tibia i fibula) su čvrsto spojene, poput zlatne mladeži (Chrysochloris); đon je presavijen prema van, kao kod Chrysochlorisa; 5. prst je savijen na plantarnu površinu, 5. nokat na stopalu je širok i lopatičan, 4. je dugačak i srpast, 3. i 2. postaju sve kraći i širi; na prsnoj kosti visoko razvijeni greben, poput zlatne krtice, krtice (Spalax) i krtice (Talpa) (sl. 549).
Snažan razvoj grebena na kostima za pričvršćivanje mišića, skraćivanje karpusa, čvrsta veza karpalije, smanjenje nekih falangi i jačanje drugih koje igraju ulogu u kopanju - svi ovi znakovi su karakteristični za sisari koji se kopaju uopšte. Ali ovisno o tome kakvo tlo životinja kopa, bit će razlike. Prilikom kopanja u mekom tlu (na primjer, u krtici - Talpa) (Sl. 550), šaka se snažno širi, djeluje kao lopata i ima tendenciju da razvije 6. prst. Prednji ud krtice je uporediv s lopatom koja se koristi za kopanje u mekom tlu. Naprotiv, kod onih oblika koji kopaju u čvrstom tlu ruka postaje uža, u kopanju učestvuje samo nekoliko prstiju, uglavnom 3. i 4., koji se razvijaju jači od ostalih, kao kod Notoryctes i Chrysochloris. Ova četka je uporediva sa podlogom za kopanje tvrdog tla.
![]()
Isto vidimo kod sisara koji kidaju čvrste strukture termita, na primjer, kod mravojeda (Myrmecophagus), kod tamandue (Tamandua). Ove životinje hodaju, oslanjajući se na vanjsku ivicu stopala, savijenih kandži.
Prilagodba sisara na brzo kretanje po tlu izazvala je vrlo značajne promjene na udovima. Kretanje po tlu, koje je bilo karakteristično za primitivne sisare, vršilo se oslanjanjem na cijelu ruku i stopalo; originalne forme sisari su prvenstveno bili plantigradni ili plantigradni, odnosno pri hodu su se oslanjali na čitava stopala, kao na primjer medvjed. Ali takva struktura dopušta samo relativno sporo hodanje u usporedbi s brzonogim antilopama ili konjima, ili skačućim zečevima, kengurima, jerboasima. Tokom filogenetskog razvoja vidimo postupno izdizanje iznad tla ručnog zgloba i tarzusa (karpus i tarsus), nakon čega su slijedili metakarpus i metatarsus, ali su falange i dalje u potpunosti dodirivale tlo. Na kraju su se i falange ispravile, prednji i stražnji udovi počeli su se oslanjati na krajeve prstiju. Sada možemo pratiti faze ove evolucije korak po korak ne samo kod kopitara, već i kod sisara mesoždera koji su stekli sposobnost brzog trčanja. Kako se udovi ispravljaju, ekstremni prsti prestaju dodirivati tlo, prestaju funkcionirati i atrofiraju. To rezultira manjom površinom kontakta sa tlom. Promjene u distalnom dijelu udova dovele su do promjena u proksimalnom dijelu: karpalija i tarsalija se nalaze sve čvršće jedna uz drugu, ulna i fibula su smanjene, udovi počinju da se kreću samo u sagitalnoj ravni, srednja metapodija se spajaju u jedna kost.
Dva razvojna puta vode do formiranja udova modernih kopitara. U jednom redu osovina ekstremiteta prolazi sredinom trećeg nožnog prsta i ovaj se najjače razvija, dok ostali postaju rudimentarni. To su kopitari (Perissodactyla), mezaksijalni (Mesaxonia). Kod drugih se osovina kreće između trećeg i četvrtog prsta, razvijajući se ravnomjerno: artiodaktili (Artiodactyla), paraksijalni (Paraxonia). Bočni prsti su smanjeni. Razvoj kopitara najjasnije se vidi u filogenetskom razvoju udova konja (Sl. 551). Razvoju papkara, između ostalog, doprinijelo je i to što mu je prethodila redukcija prvog prsta, tako da u prelasku sa hodanja stopala gotovo da nije učestvovao, dok kod predaka kod jednopapka, razvoj petog prsta je u početku bio normalan.
Zanimljivo je da su se i artiodaktil i artiodaktilar pojavljivali u evolucijskom procesu više puta i u nekim slučajevima doveli do vrlo specijaliziranog tipa, na primjer, kod južnoameričkih Proterotlieriidae (izumrli).
Preostali prsti su obloženi kopitima, koji se mogu dodatno razlikovati ovisno o načinu života i staništu. Na primjer, koze koje se drže za kamenje imaju strma i uska kopita sa oštrim rubom; at irvasi veoma su dugačke, široko račvane i služe kao neka vrsta skija koje sprečavaju životinju da padne u sneg i močvare.
Istovremeno sa prstima, sa razvojem hodanja na prstima, dolazi do promjene u zglobu (carpus) i tarsusu (tarsus). To ide drugačije kod artiodaktila i kopitara zbog različite distribucije gravitacije. Konačno, u vezi s prelaskom na nalcewalking, vrijednost lakatne kosti (ulna) kao oslonca za rame postepeno se smanjuje: smanjuje se ili se spaja sa radijusom (radijusom). Isto tako, fibula se smanjuje i konačno nestaje u zadnjim udovima.
Značenje i priroda opisanih promjena jasno su vidljivi iz poređenja udova kamile i slona (sl. 552). Kod slona šaka (stopalo) ostaje kratka, a rame i podlaktica (bedro i potkolenica) su produženi. Naprotiv, kod kamile su narasle metapodije produžene; nadlaktica i podlaktica (kao i butina i potkolenica) ostaju kratke. Zbog gubitka određenog broja kostiju, skelet ekstremiteta je pojednostavljen, potonji postaje lakši. Smanjenje prstiju dovodi do manjeg trenja, smanjuje se rizik od ozljeda, rupture ligamenata. Fuzija metapodija stvara veliku snagu. Ud slona je mnogo teži. Obilje kostiju u ruci zahtijeva spajanje svih prstiju kako bi se postigla stabilnost. opšta koža... Ud slona je stub velike snage, ali mu nedostaje lakoća koju ima ud kamile.
Drugi način da se brzo krećete po zemlji je skakanje. Ova se adaptacija više puta pojavljivala u istoriji sisara i, osim toga, u širokom spektru filogenetskih stabala: među tobolčarima - u kengura (Macropus), među insektivorima - među skakačima (Macroscelidae) i, konačno, u različiti činovi razvoj glodara: ovo. zečevi (Leporidae) iz Duplicidentata; u seriji dikobraza (Hystricomorplia) razvila se grupa šetača (Pedetinae); među mišolikim vrstama (Myomorpha) imamo australskog jerboa (Conilurus), jerboa iz Starog svijeta (Jaculus, Alactaga), američke skakače (Zapus) i mrežaste skakače (Dipodomys).
Razvoj sposobnosti skakanja povezan je sa produžavanjem zadnjeg ekstremiteta, odnosno kostiju metatarzusa (metatarsus). Osim toga, to dovodi do gubitka ekstra ekstremnih prstiju, a kosti tarzusa su međusobno vrlo čvrsto povezane. Primjer su noge zemljanog zeca (Alactaga) i pravog jerboa (Dipus).
Alactagine prednje noge su male i kratke. Zemljani zec ih koristi prilikom kopanja, ali ne dodiruje zemlju kada skače. Skakanje se izvodi samo uz pomoć stražnjih nogu, pa čak i bez pomoći repa. Potonji dodiruje tlo samo u mirnom položaju, kada se životinja oslanja na rep. Skelet noge ima osobinu da se metatarzalne kosti 2., 3. i 4. prsta spajaju u jednu kost. Krajevi metatarzalnih kostiju ispod nisu spojeni, a medijan (3. prst) strši dalje naprijed. Od falangi povezanih s njima, srednje su opet najduže. Prvi i peti prst su rudimentarni i ne dodiruju tlo. Oba ova prsta imaju jednake dužine zbog činjenice da metatarzalna kost thumb duže od 5. Prvi ima 2, drugi - 3 falange.
U Dipusu je specijalizacija otišla još dalje: 1. prst je netragom nestao, od 5. je ostao samo rudiment. Srednji prst duplo slabiji od dva ekstremna, a tek nešto duži od njih. Nožni prsti su paralelni. Srednji prst je smanjen jer njegovu funkciju preuzimaju 2. i 4. na čijim krajevima se nalaze naglasci pri sjedenju i skakanju. U daljoj evoluciji, treći prst bi trebao nestati: imamo nedovršenu vrstu adaptacije u Dipusu.