Quelle est la colonne vertébrale thoracique des mammifères. Structure externe des mammifères, squelette et muscles sur l'exemple d'un chien domestique
La colonne vertébrale comprend les régions cervicale, thoracique, lombaire, sacrée et caudale. Le sien caractéristique- forme platycéliale (avec des surfaces planes) des vertèbres, entre lesquelles il y a des cartilagineux disques intervertébraux... Les arcades supérieures sont bien définies. Dans la région cervicale, il y a sept vertèbres, dont la longueur dépend de la longueur du cou; seulement chez le lamantin et le paresseux - Choloepus hof-fmani, il y en a 6, et chez le paresseux - Bradypus 8-10. Les vertèbres cervicales sont très longues chez la girafe et très courtes chez les cétacés qui n'ont pas d'interception cervicale. Les côtes qui forment la poitrine sont attachées aux vertèbres de la région thoracique. Le sternum le fermant est plat et seulement à chauves-souris et les espèces fouisseuses avec des membres antérieurs puissants (par exemple, les taupes) ont une petite crête (quille) servant de point d'attache pour les muscles pectoraux. Dans la région thoracique, il y a 9-24 (généralement 12-15) vertèbres, les 2-5 dernières vertèbres thoraciques portent - de fausses côtes - n'atteignant pas le sternum., Dans la région lombaire de 2 à 9 vertèbres; les côtes rudimentaires se confondent avec leurs grandes apophyses transverses. La région sacrée est formée de 4 à 10 vertèbres fusionnées, dont seules les deux premières sont vraiment sacrées et les autres sont caudales. Le nombre de vertèbres caudales libres varie de 3 (chez le gibbon) à 49 chez le dinosaure à longue queue.
Le degré de mobilité des vertèbres individuelles est différent. Chez les petits animaux qui courent et grimpent, il est grand, sur toute la longueur de la colonne vertébrale, de sorte que leur corps peut se plier en différentes directions et même se rouler en boule. Les vertèbres des régions thoracique et lombaire sont moins mobiles chez les grands animaux qui se déplacent rapidement. Chez les mammifères voyageant sur pattes postérieures ah (kangourous, gerboises, sauteurs), les plus grosses vertèbres sont situées à la base de la queue et du sacrum, et plus loin leur taille diminue progressivement. Chez les ongulés, au contraire, les vertèbres et surtout leurs apophyses épineuses sont plus grosses dans la partie antérieure de la région thoracique, où s'attachent les puissants muscles du cou et en partie les membres antérieurs (
La structure externe du chien
Le chien a été le premier animal de compagnie. L'homme l'a apprivoisée dans les temps anciens. Le chien aidait l'homme primitif pendant la chasse, gardait sa maison. Les chiens de service, de chasse et de décoration, ainsi que les bâtards (bâtard) sont désormais connus. Les chiens aident à effectuer le service pour protéger les frontières de notre patrie, et pendant la Grande Guerre patriotique recherché les blessés, aidé dans la reconnaissance et les communications. Sur les chiens, les scientifiques mènent des études sur l'activité vitale du corps. Chien avant l'homme espace visité.
Le corps d'un chien, comme un berger ou un husky, est mince et musclé. Ses membres ne sont pas situés sur les côtés du corps, comme chez les reptiles, mais sous le corps, de sorte que le corps de l'animal ne touche pas le sol. Le chien marche appuyé sur des doigts avec de fortes griffes. Un cou flexible favorise une grande mobilité de la tête. La bouche de l'animal est limitée par des lèvres mobiles - supérieure et inférieure. Au dessus la lèvre supérieure il y a un nez avec une paire d'ouvertures nasales externes - les narines. Les yeux ont des paupières bien développées. La membrane clignotante (troisième paupière) chez un chien, comme chez tous les mammifères, est sous-développée. De tous les animaux, seuls les mammifères ont l'oreille externe - Auricule... L'oreille externe du chien est grande et mobile.
Couvertures de mammifères
La peau des mammifères est solide et élastique; chez la plupart des animaux, les bases des poils y sont placées, ce qui constitue la caractéristique de cette classe de vertébrés Racine des cheveux ... Distinguer entre épais et cheveux longs — la colonne vertébrale et plus court, plus doux - sous-sol, ou couche... L'aube grossière et durable protège le sous-poil et la peau des dommages. Le sous-poil, qui retient beaucoup d'air, retient bien la chaleur corporelle. En plus de l'arête et du sous-poil, les animaux développent cheveux larges- les organes du toucher.
Les poils des mammifères, comme les plumes des oiseaux et les écailles des reptiles, sont constitués de matière cornée. Deux fois par an, le chien, comme de nombreux autres mammifères, perd - certains de ses poils tombent et sont remplacés par de nouveaux. La base des cheveux est située à l'intérieur d'un sac à cheveux spécial, où les conduits du voisin s'ouvrent glandes sébacées... Leurs sécrétions lubrifient la peau et les cheveux, ce qui les rend élastiques et non mouillés par l'eau.
Motif : La structure de la peau et des poils d'un mammifère
Dans la peau de la plupart des animaux se trouvent et glandes sudoripares ... La sueur s'évapore de la surface du corps et le refroidit. Avec la sueur, l'excès de sel et d'urée est également excrété par le corps; ainsi, les glandes sudoripares jouent un rôle corps supplémentaires décharge.
Le chien glandes sudoripares peu et le refroidissement du corps est obtenu avec une respiration accrue.
Aux extrémités des orteils des mammifères se trouvent des griffes, des ongles ou des sabots cornés. Parfois, des formations cornées se développent également sur la tête (cornes chez les rhinocéros, les antilopes, les bovins, etc.) ou sur la queue (par exemple, les écailles cornées chez les rats).
Squelette de mammifère
Le squelette des mammifères se compose des mêmes sections que chez les autres vertébrés. Le crâne des animaux se distingue par un crâne plus grand, qui est associé à grande taille cerveau. Pour les mammifères, la présence de 7 vertèbres cervicales est très caractéristique. Les girafes à long cou et les baleines ont le même nombre de vertèbres cervicales. Les vertèbres thoraciques (généralement 12-15), ainsi que les côtes et le sternum, forment une poitrine solide. Vertèbres massives lombaire articulés les uns avec les autres de manière mobile. Dans cette section, le corps peut se plier et se déplier. Le nombre de vertèbres lombaires est différent selon différents types(2-9), le chien en a 6. L'épine sacrée (3-4 vertèbres) fusionne avec les os du bassin. Le nombre de vertèbres caudales (de trois à plusieurs dizaines) dépend de la longueur de la queue.

Figure : Le squelette de mammifères à l'aide de l'exemple d'un chien

Motif: Crâne de chien
Ceinture des membres antérieurs mammifères se compose de deux omoplates auxquelles sont attachés des os de corbeau et de deux clavicules. Les clavicules du chien ne sont pas développées. Ceinture de membre postérieur- bassin - formé de trois paires os pelviens... Les squelettes des membres chez différentes espèces de mammifères et de reptiles sont similaires, cependant, les détails de leur structure chez différentes espèces sont différents et dépendent des conditions de vie de l'animal.
Muscles de mammifères
La plupart des mammifères, y compris le chien, ont particulièrement développé les muscles du dos, des membres et de leurs ceintures. Le chien peut courir à grands pas, plier et redresser le corps, poussant alternativement du sol avec ses pattes avant ou arrière. Des muscles forts déplacent la mâchoire inférieure. En serrant les mâchoires, armé de dents, le chien tient fermement sa proie.

Motif : Muscles du chien déplaçant la mâchoire inférieure
Ceinture d'épaule mammifères est simplifiée, se compose d'un grand omoplates ayant une excroissance coracoïde, et clavicule(Fig. 80, 81). Au stade de développement embryonnaire et chez les monotrèmes adultes coracoïde est un os indépendant.
Omoplate situé en haut poitrine, a une forme triangulaire et une excroissance élevée (crête) avec acromial un processus pour l'attachement des muscles. À l'emplacement de l'excroissance coracoïde et du processus acromial de l'omoplate, il y a fosse glénoïde pour attacher le membre antérieur.
Clavicule en forme de tige sert à relier l'épaule et le sternum. La clavicule est plus développée chez les animaux effectuant les membres antérieurs. mouvements complexes(chauves-souris, taupes, primates, rongeurs, etc.). Chez de nombreuses espèces, caractérisées par des mouvements monotones (ongulés, chiens), la clavicule est absente. En général, la ceinture des membres antérieurs est reliée au squelette axial par des muscles et des ligaments.
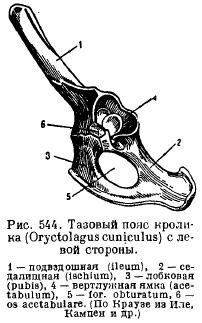
Ceinture pelvienne (fig. 82 A) se compose de paires os sans nom articulé iliaque, sciatique et pubien OS. Bassin type fermé et à travers l'ilion est fermement connecté à sacré la colonne vertébrale. Lorsque les éléments du bassin grandissent ensemble, union pubienne et formé cotyle pour la fixation des membres postérieurs. Dans la partie inférieure de l'os de l'anneau, il y a trou obturateur.
Squelette de membres appariés
Les membres des mammifères, comme ceux des autres vertébrés terrestres, sont du type à cinq doigts et représentent un levier à trois membres, dont toutes les parties sont interconnectées de manière mobile.
Membre antérieur consiste en épaule, avant-bras et main(fig. 81). Os brachial bien développé, a une tête arrondie pour la fixation à la ceinture du membre. Avant bras faire deux os - radial et ulnaire ayant olécrâne au sommet. A la jonction de l'épaule et de l'avant-bras se trouve articulation du coude.
Brosser comprend trois départements - poignet, métacarpe et phalanges. Poignet se compose de 8-10 éléments disposés en trois rangées ... Paturon comprend cinq os (selon le nombre de doigts). Chaque orteil a trois phalanges, à l'exception du premier orteil, qui a deux phalanges. Contrairement à l'articulation intercarpienne des reptiles et des oiseaux, le mobile découper chez les mammifères situés entre l'avant-bras et la main(comme les amphibiens).
membre postérieur mammifères a également trois divisions - cuisse, bas de jambe et pied(Fig. 82B). Fémur massive et insérée avec une tête arrondie dans le cotyle de la ceinture pelvienne. Le tibia comprend deux os - tibia et tibia. Le plus grand tibia est l'élément interne du tibia et le péroné est sa partie externe.
Entre la cuisse et le bas de la jambe, il y a découper couvert genouillère qui ont évolué à partir de leurs tendons musculaires ossifiés. Les surfaces des os articulés sont recouvertes de formations cartilagineuses. Articulation de la cheville mammifères est situé entre les extrémités inférieures des tibias et la partie proximale du pied.

Riz. 82. Ceinture pelvienne (A) et membre postérieur (B) du renard :
1 – ilion, 2 - ischion, 3 - os pubien, 4 - acétabulum, 5 - fémur, 6 - rotule, 7 - tibia, 8 - péroné, 9 - tarse, 10 - métatarse, 11 - phalanges des doigts, 12 - talus, 13 - calcanéum
Éléments du tarse les pieds sont disposés en trois rangées et forment une excroissance - calcanéum typique des mammifères. Métatarse se compose de cinq os, auxquels les phalanges des doigts sont attachées. Comme dans les membres antérieurs, il y en a trois dans chaque orteil, à l'exception du premier (intérieur), qui n'a que deux phalanges.
Pour les mammifères, un positionnement particulier des membres par rapport au corps est caractéristique : ils sont situés sous le corps, ce qui contribue au développement d'une plus grande maniabilité des mouvements. Dans ce cas, l'articulation du coude est dirigée vers l'arrière, et articulation du genou au contraire, il est orienté en avant.
En relation avec la diversité des espèces des mammifères, leur mode de vie et leurs modes de déplacement, la longueur relative des membres, le rapport des éléments constitutifs, la configuration des os, le nombre de doigts, etc., varient considérablement. chez les chauves-souris, la longueur du membre antérieur, en particulier les phalanges des doigts, servant à attacher la membrane aérienne.
Au contraire, le membre antérieur de la taupe est raccourci, la partie palmaire de la main est élargie et le sixième orteil est bien développé. Chez les baleines, la main est fortement allongée en raison de l'augmentation du nombre de phalanges des doigts, à la suite de laquelle le membre a acquis grande similitude avec une nageoire de poisson à nageoires croisées. Les doigts des animaux à grande biomasse et à course rapide (équidés, artiodactyles) ont été significativement réduits et modifiés.
L'une des caractéristiques du squelette des mammifères est grand développement cartilage chez les jeunes animaux et sa préservation à long terme dans la période de développement post-embryonnaire. Cela est dû aux particularités de la croissance du cartilage et des os en relation avec la croissance de l'animal lui-même. C'est le cartilage qui croît avec toute sa masse (interstitielle), la croissance se produit rapidement et les proportions des parties peuvent être préservées, tandis que l'os croît plus lentement et uniquement à partir de la surface, de sorte que les parties du squelette qui apparaissent toujours restent à la même distance et la préservation de la proportion de pièces avec la croissance de l'os s'avère impossible. C'est pourquoi les jeunes animaux contiennent toujours beaucoup de cartilage, et surtout dans les endroits où se déroule la croissance. Alors, os longs les membres sont toujours constitués d'une partie médiane - la diaphyse, qui s'ossifie tout d'abord, et les extrémités, qui sont longtemps conservées par le cartilagineux - la glande pinéale. L'ossification de la diaphyse commence tout d'abord à partir de la périphérie - périchondrale, c'est-à-dire due aux cellules provenant du tissu conjonctif qui recouvre le cartilage - à partir du périchondre; puis l'ossification interne du cartilage le rejoint - ossification endochondrale; ce n'est que plus tard, lorsque des cavités médullaires apparaissent dans la diaphyse par désintégration de l'os, que les glandes pinéales commencent à s'ossifier de manière endochondrique; entre la diaphyse et les épiphyses, cependant, il reste longtemps du cartilage, à l'aide duquel l'os s'allonge en conservant les proportions (fig. 530).
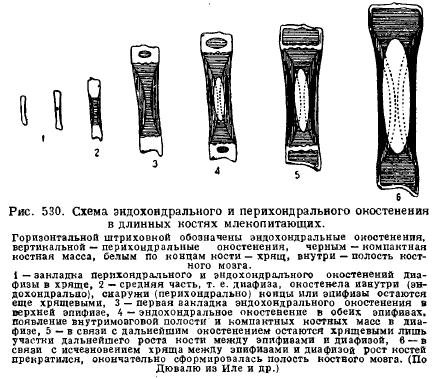
Colonne vertébrale les mammifères se caractérisent, contrairement à Sauropsida, par l'absence d'articulations libres entre les vertèbres, à l'exception de la première et de la deuxième vertèbre. Les disques de tissu conjonctif élastique intervertébral se situent entre les corps vertébraux. En général, cela se traduit par moins de flexibilité de la colonne vertébrale, ce qui est dû au mouvement plus spécialisé des mammifères à l'aide des membres.
L'ossification des corps vertébraux se produit par la glande pinéale cartilagineuse. Les corps ont des surfaces planes ou légèrement convexes, chez la plupart des ongulés modernes, ils sont légèrement opisthocèles. Sur les vertèbres, les apophyses épineuses sont bien développées, notamment dans la région thoracique, ainsi que les apophyses transverses qui siègent sur des arcs ; à la base de ce dernier il y a des processus articulaires.

Le nombre de vertèbres cervicales est généralement de 7 (Fig. 531), quelle que soit la longueur du cou, de sorte que la longueur du cou, contrairement aux oiseaux, dépend uniquement de la longueur des vertèbres. Seuls le lamantin (Manatus) et une espèce de paresseux (Choloepus hoffmani) possèdent 6 vertèbres cervicales, tandis que les paresseux à trois doigts (Bradypus) en possèdent 9. Le cou des cétacés, qui a failli disparaître à cause du mode de vie aquatique, en contient encore 7 , fusionnant partiellement les vertèbres, alors que dans le cou d'une girafe, ces 7 vertèbres sont très longues. Les 2 premières vertèbres cervicales sont modifiées en un atlas et une épistrophie, reliées par de vraies articulations, et chez les monotrèmes, le processus odontoïde fonctionne toujours sous la forme d'un os spécial, et n'a pas fusionné, comme chez d'autres mammifères, avec le corps de l'épistrophie. Les côtes rudimentaires de la région cervicale se développent avec les vertèbres, ce qui entraîne la formation d'apophyses transverses perforées. La fusion partielle ou complète des vertèbres cervicales se produit chez les cétacés; dans les gerboises, toutes les vertèbres cervicales se développent ensemble à l'exception de la 7e, dans la taupe marsupiale (Notoryctes) elles fusionnent de la 2e à la 6e, dans la taupe - de la 3e à la 4e.
Le nombre de vertèbres thoraciques, caractérisées par des côtes qui leur sont attachées, varie de 9 [chez un cétacé (Hyperoodon) et un tatou (Tatusia)] à 25 [chez un paresseux], mais il y en a généralement 12-13. Les processus sur les vertèbres thoraciques sont bien développés : épineux (proc. Spinosus), transverses ou diapophysaires (proc. Transversus), articulaires ou zygapophyses (proc. Articularis) ; les côtes sont reliées par leur tête au corps d'une ou deux vertèbres adjacentes et par des tubercules aux apophyses transverses. Ils se composent de deux parties: côte osseuse - proximale et cartilagineuse - distale. Seules les côtes des vertèbres thoraciques antérieures atteignent le sternum, les côtes postérieures se terminent librement (fausses côtes). Les vertèbres lombaires, qui se caractérisent par l'absence de côtes incluses ici dans la composition des apophyses transverses, chez les mammifères de 2 à 9, généralement 6-7.
Les vertèbres sacrées des mammifères sont les vertèbres qui, ayant grandi ensemble, sont articulées avec l'ilion... Il existe 1 à 10 de ces vertèbres, mais seulement 1 à 2 d'entre elles sont de véritables vertèbres sacrées, c'est-à-dire celles dans lesquelles l'articulation avec le bassin se fait par des ossifications indépendantes correspondant aux côtes sacrées; le reste des vertèbres sacrées sont fausses, c'est-à-dire les vertèbres caudales attachées au sacrum.
Le nombre de vertèbres incluses dans la région caudale varie énormément : de 3 ( les chauves-souris, gibbon) à 49 [lézard (Manis macrura)].
Sternum(sternum) des mammifères se développe à partir de la fusion des extrémités ventrales des côtes, qui forment alors une plaque non appariée, composée d'un certain nombre de départements (Fig. 532).
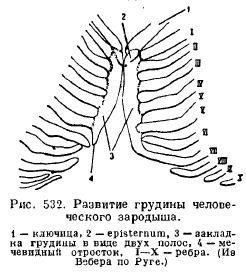
Relié aux clavicules (le cas échéant) et à la première paire de côtes, le sternum antérieur étendu s'appelle la poignée (praesternum, manubrium); suivi d'un département appelé le corps (mesosternum, corpus), souvent divisé en une série d'os, situés séquentiellement entre les paires de côtes suivantes ; encore plus loin se trouve le processus dit xiphoïde (xiphisternum). Toutes les côtes ne sont pas en rapport avec le sternum, un nombre plus ou moins important de fausses côtes se termine librement (Fig. 533).

Chez les mammifères cloacaux (Monotremata), en raison de la présence de coracoïdes, absents chez tous les autres mammifères, devant l'anse du sternum, le cartilage du pré-sternum (prosternum) est posé, ce qui est en rapport avec le coracoïdes, qui est ensuite déplacé par l'os en développement - l'épisternum (episternmn), qui rapproche le cartilage cloacal des reptiles.
Godille les mammifères diffèrent par un certain nombre de caractéristiques du crâne des reptiles. Cependant, ces caractéristiques ne sont pas apparues immédiatement, mais à travers un changement progressif, qui peut être rencontré en étudiant les crânes de bêtes lézard fossiles (Theromorpha), en particulier de l'ordre des animaux à dents animales (Theriodontia). Crâne de mammifères - le crâne est tropibasal, mais avec une partie interorbitale raccourcie, à cause de laquelle il ressemble à un platybasal, avec un grand crâne, avec une tendance prononcée à la fusion des os individuels les uns avec les autres,
L'emplacement des os dans le crâne des mammifères peut être jugé par le schéma ci-joint (Fig. 534).
Sur les os occipitaux latéraux, à l'extérieur des tubercules articulaires, il existe généralement un processus occipital (pp. Paroccipitalis), qui est particulièrement bien développé chez les rongeurs et chez de nombreux ongulés. Devant les tubercules articulaires, il y a généralement une ouverture à l'extérieur du nerf hyoïde (XII) (pour. Condyloideum anterius).
Plusieurs ossifications sont déposées dans le cartilage de la capsule auditive, qui, cependant, se confond très tôt en deux os: couché devant - pierreux (pétrosum) et couché à l'extérieur et derrière - mastoïde (mastoideum); ces deux os, fusionnés, donnent l'os pétro-mastoïdien, ou simplement pierreux (pétromastoideum, pétrosum ou périotique), renfermant le labyrinthe auditif interne. Bordures du périotique dans la partie postérieure avec les os occipitaux principaux et latéraux, et dans la partie antérieure avec les os sphénoïde et aile-sphénoïde ; puisque des vaisseaux et des nerfs passent entre ces os et l'os pierreux, des lacunes importantes subsistent ici : derrière - l'ouverture lacérée postérieure (pour lacerum posterius), ou l'ouverture jugulaire (pour jugulare) pour le passage du nerf glossopharyngien (IX), branches du nerf vague ( X) et pour le nerf accessoire (XI), ainsi que pour la veine jugulaire interne, et en avant - le trou de lacération antérieur (pour. Lacerum anterius), à travers lequel l'artère carotide interne passe dans le crâne . Parfois, la connexion de l'os pétreux avec le crâne est si faible qu'il peut facilement être séparé avec l'os tympanique qui y adhère, ou même lorsque le crâne est macéré, il tombe complètement, comme c'est le cas chez les cétacés.
En dehors de la région de l'oreille, deux os tégumentaires sont formés. L'un d'eux, dont l'origine n'est pas encore suffisamment élucidée, se développe au niveau du conduit auditif externe sous la forme d'un petit os semi-circulaire. Il s'agit de l'os tympanique (tympanum), qui est probablement un homologue de l'os angulaire (angulare) mâchoire inférieure reptiles qui se sont déplacés dans la région parotide avec d'autres os de la mâchoire inférieure en raison de changements dans l'articulation de la mâchoire. Situé initialement sous la forme d'un os semi-circulaire ou annulaire dans la zone du conduit auditif, il se développe souvent et forme, généralement avec l'os pierreux et parfois écailleux, le méat auditif externe (meatus auditorius externus). La membrane tympanique est tendue sur l'os tympanique. Plus profondément vers l'intérieur, l'os tympanique peut également être impliqué dans la formation des parois de l'oreille moyenne. Souvent, l'os tympanique sur la face inférieure de la région auditive se gonfle en une bulle à paroi mince (bulla ossea).
Le deuxième os d'origine cutanée, qui participe à la formation du squelette de la région de l'oreille, est l'os squameux (squamosum) ; il a reçu un développement et une importance exceptionnels chez les mammifères en rapport avec l'attachement de la mâchoire inférieure à celui-ci. Chez les singes et les humains, tous ces os de la région de l'oreille se développent ensemble pour former un complexe temporal ; l'os squameux n'en constitue ici qu'une partie, à savoir les écailles (squama temporalis) os temporal, tandis que l'os tympanique est la partie tympanique (pars tympanicum), et les os cartilagineux parotides eux-mêmes forment les parties de la pierre (pars petrosa) et de la mastoïde (pars mastoidea). Le degré de développement de l'os squameux et sa participation à la formation des parois de la boîte cérébrale et du méat auditif sont très différents, mais l'os squameux a toujours une surface articulaire (cavitas glenoidea) pour l'articulation de la mâchoire inférieure. Ainsi, les mammifères, contrairement à tous les autres vertébrés, ont une attache originale très particulière de la mâchoire inférieure : non pas à l'os carré, mais à l'os squameux du crâne. La surface articulaire de la mâchoire inférieure repose sur la base s'étendant des écailles vers l'extérieur et l'avant processus zygomatique(proc. zygomaticus). Dans les monotrèmes au-dessus de la surface articulaire, il est pénétré par le canal temporal (can.temporalis), qui est particulièrement prononcé dans l'ornithorynque. Derrière la fosse articulaire, il y a un processus (proc. Postglenoideus).
Devant la partie auditive, la base du crâne est formée par l'os sphénoïde principal (basisphenoidale), sur surface intérieure qui présente une dépression plus ou moins importante - la selle dite turque (sella turtica), dans laquelle se trouve l'hypophyse. Sur les côtés de l'os sphénoïde principal se trouvent, formant les parties inférieures de la paroi antérieure du crâne, des os alaires sphénoïdes (alisphenoidea), généralement fusionnés avec lui en un seul os, formant de grandes ailes (alae magnae) de cet os. Les os aile-sphénoïde sont munis de deux trous : un ovale (for.ovale) pour le passage de la troisième branche du nerf trijumeau (V3) et rond (for.rotundum) pour le passage de la deuxième branche du même nerf (V2). Cependant, le premier se confond souvent avec l'ouverture déchiquetée antérieure (pour. Lacerum anteris), et le second se confond souvent avec l'ouverture orbitale en coin (pour. Sphenorbitale), qui est simultanément délimitée par l'os sphénoïde et l'os orbital- l'os sphénoïde se trouvant devant lui. Par ce trou passent les nerfs de l'oculomotorius III, du trochlearis IV, de l'abducens VI et de la première branche du nerf trijumeau.
La couverture crânienne est ici recouverte d'os pariétaux (pariétal), qui grandissent souvent ensemble (chez les monotrèmes, certains marsupiaux et ongulés); entre eux et l'os occipital supérieur, deux autres os sont posés, fusionnant en un os interpariétal (interparietale), qui dans certains ordres est conservé comme un os indépendant (cétacés, marsupiaux, hyrax, oryctéropes, ailes laineuses et certains rongeurs), tandis que dans d'autres, il pousse ensemble ou avec le pariétal (chez certains rongeurs, sirènes et ruminants), ou avec l'os occipital supérieur (chez les primates carnivores, à un sabot).
Devant l'os sphénoïde principal se trouve l'os sphénoïde antérieur (praesphenoideum) avec une dépression pour le chiasma des nerfs orbitaires; sur ses côtés, généralement fusionnés avec lui, se trouvent les os orbitaux-sphénoïdes (orbitosphenoidea) avec une ouverture pour le passage des nerfs orbitaires (pour. opticum), appelés en cas de fusion avec les os sphénoïdes antérieur et principal, petites ailes (alae parvae). Comme mentionné ci-dessus, entre les deux ptérygoïdes de chaque côté, il y a des ouvertures en forme de fente à travers lesquelles passent les nerfs des muscles oculaires et la première branche du nerf trijumeau (pour. Sphenorbitale).
Au sommet de cette rangée d'os correspondent les os frontaux tégumentaires (frontaux) - des os appariés généralement bien développés, parfois fusionnés et portant des tiges en cornes de bovin, et en cerf - chanvre pour les bois. Couvrant le crâne, les os frontaux sont limités d'en haut et de l'intérieur de l'orbite, formant souvent des processus : os zygomatique(jugale) avec l'os frontal, l'anneau périobital (certains insectivores et carnivores, ruminants, unonglos, semi-singes). Chez les singes, le processus de séparation de l'orbite de l'orbite temporale va si loin qu'une paroi osseuse se forme entre l'orbite et l'orbite temporale.
Dans la paroi antérieure de l'orbite se trouve un petit os lacrymal tégumentaire (lacrymale), perforé par l'ouverture du canal lacrymal (pour. Lacrymale).
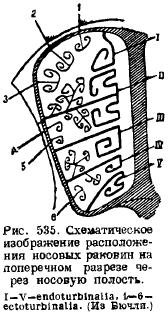
Dans la région nasale, plusieurs ossifications sont posées dans la capsule nasale cartilagineuse. Tout d'abord, le long de la cloison nasale cartilagineuse, une cloison osseuse (lame perpendiculaire) se développe au milieu, correspondant à l'os olfactif moyen (mésethmoïde); deuxièmement, dans mur arrière la capsule nasale et dans la paroi antérieure de la cavité cérébrale, se développent deux plaques latérales, perforées par de nombreux [seulement dans l'ornithorynque ces trous sont remplacés par un (pour. olfactorium)] trous pour le passage du nerf olfactif - plaques ethmoïdales ( lamina cribrosa), correspondant aux os ethmoïdaux latéraux (ethmoidalia lateralia) ... À partir de la cloison nasale, l'ossification s'étend de la face dorsale le long des côtés, longe les parois latérales des capsules nasales et se termine au fond de celles-ci, formant de nombreuses protubérances sur les côtés à l'intérieur de la cavité nasale. Ce sont des coquilles olfactives (ethmoturbinales) qui, avec les plaques latérales et les cavités d'une structure cellulaire, forment parfois un labyrinthe nasal très complexe. La position des éviers peut être vue sur le schéma ci-joint (Fig. 535). De l'os nasal tégumentaire (nasale), qui couvre cette zone d'en haut, pendent à l'intérieur de la cavité nasale, également une de chaque côté, des coquilles (nasoturbmalia). Bas sur cloison nasale l'os non apparié, qui était auparavant considéré comme un vomer, se chevauche. Cependant, des études récentes ont montré qu'en plus de cet os tégumentaire non apparié, deux os appariés, appelés praevomer, se développent à la base du crâne ; et puisque les reptiles ont un vomer sous la forme d'un os apparié, et qu'un parasphénoïde était un os non apparié dans cette zone, c'est le vomer des mammifères qui peut être considéré comme un homologue de ce dernier os des reptiles, et les pré-vomers sont considérés comme des vomers. À l'avant, les parois de la cavité nasale sont les os de la mâchoire (maxillaire), sur lesquels se forment des coquilles qui se développent dans la cavité nasale; ce sont des coquilles de mâchoire (maxilloturbinale).
La face antérieure du crâne se termine devant les os intermaxillaires (praemaxillare). Chez les singes anthropomorphes et chez l'homme, cet os se développe très tôt avec la mâchoire, et chez les chauves-souris il est réduit à un petit os ou même totalement absent. Avec son processus vertical, l'os intermaxillaire limite l'avant de l'ouverture nasale et s'étend entre l'os nasal et la mâchoire très en arrière, parfois jusqu'à l'os frontal. D'en bas, les os intermaxillaires participent à la formation d'un palais dur. Ils limitent l'avant et les côtés sur le palais dur des ouvertures du canal palatin-posovy (stenson) (pour. Incisiva), reliant cavité buccale avec un orgue Jacobson. Les processus des os intermaxillaires situés à l'extérieur de ces ouvertures palatines-nasales sont superposés pour chaque os indépendamment ; ils sont probablement des homologues du vomer apparié de reptiles (praevomer) et sont conservés dans l'ornithorynque en tant qu'os indépendants. Les incisives sont situées sur l'os intermaxillaire.
Les os de la mâchoire (maxillaire), comme déjà mentionné, limitent le fond de la cavité nasale, formant un palais dur avec des processus horizontaux. Sur la face interne de la cavité nasale, la coquille portable inférieure (maxilloturbines) se développe jusqu'aux parois latérales de celle-ci, formées par les os de la mâchoire, qui sont posés comme un os indépendant.
La continuation du palais dur vers l'arrière correspond aux processus horizontaux des os palatins, limitant le passage nasal interne en tant que parois latérales. Ils se tiennent verticalement et des os ptérygoïdes debout verticalement (pterygoideum) les jouxtent également par derrière, qui chez l'homme fusionnent avec l'os sphénoïde, formant ses processus ptérygoïdes (proc. Pterygoidei).
La partie antérieure du crâne subit des modifications importantes en lien avec le passage à un mode de vie aquatique chez les cétacés, les sirènes, et en lien avec le développement de la trompe chez les éléphants.

Chez les cétacés (Fig. 536), les narines sont repoussées loin dans la région interorbitaire et débouchent respectivement dans un canal vertical, s'ouvrant par les choans dans la cavité buccale ; les os nasaux se trouvent derrière les narines, chevauchant les os frontaux et presque adjacents à l'os pariétal; aussi les os de la mâchoire chevauchent les os frontaux, atteignant presque les os pariétaux, qui sont fortement décalés sur les côtés par l'os inter-pariétal.
Les sirènes ont le même recul des narines, mais pas pour autant ; les os nasaux sont encore plus réduits ici et peuvent même être absents (Fig. 537).

Chez les éléphants avec un développement exorbitant de défenses et de molaires puissantes et du fait de la présence d'une trompe, la emplacement normal les os : les narines sont fortement repoussées, à cet égard, le canal nasal prend une position presque perpendiculaire ; les os frontaux sont très courts, les os intermaxillaires sont très gros et remontent loin ; les os de la mâchoire sont déplacés sous la base du crâne ; l'ensemble du crâne devient très massif avec des os pneumatiques extrêmement développés (Fig. 538).

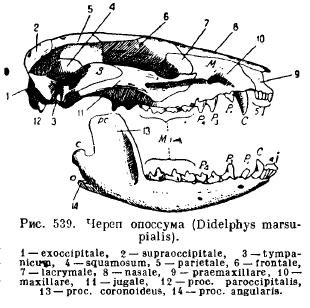
La mâchoire inférieure des mammifères, contrairement à tous les autres vertébrés, se développe de chaque côté à partir d'un seul os dentaire (dentaire); les deux moitiés restent reliées l'une à l'autre au moyen de cartilage, de couture ou grandissent ensemble (animaux à un seul sabot, éléphants, chauves-souris, singes). Chaque moitié se compose d'un corps horizontal (corpus), sur lequel reposent les dents, et d'une partie ascendante avec des processus : le processus coronal antérieur (ррос. Coronoideus), le processus articulaire situé derrière (ррос. Articularis) et chez les mammifères inférieurs, le processus angulaire en marche arrière (ррос . angularis); chez la plupart des marsupiaux et chez certains rongeurs, ce processus se transforme en une plaque s'étendant vers l'intérieur (Fig. 539).
Une telle structure de la mâchoire inférieure et son attachement particulier au crâne nous amènent à la question de l'origine de ces caractéristiques chez les mammifères. Récit développement individuel et les données paléontologiques apportent une réponse à cette question. Dans les embryons de mammifères, les cartilages palatin-carré et meckélien sont déposés. Cependant, l'ossification du cartilage carré palatin se produit en dehors de la connexion avec l'articulation de la mâchoire inférieure et ne conduit pas à la formation d'un os carré, comme chez les reptiles, et donnera un nouvel os auditif - l'enclume, tandis que le cartilage de Meckel , ossifié dans sa partie postérieure, donne un os non articulaire (articulare), comme dans les autres maxillaires, et le troisième os auditif est le marteau (malleus). Le premier osselet auditif, l'étrier, correspond à la colonne des amphibiens, des reptiles et des oiseaux et, comme nous le savons déjà, est une suspension modifiée (hyomandibulaire), c'est-à-dire la partie proximale de l'arc hyoïde. Comme la colonne (columella auris), l'étrier couvre la fenêtre ovale du labyrinthe auditif, et est perforé, comme chez certains reptiles, d'un trou à travers lequel, chez certains mammifères inférieurs, l'artère faciale (a.facialis) passe (Fig. 540 et 541).

Ainsi, chez les mammifères, l'os carré s'est transformé en enclume, l'os articulaire en marteau ; et comme un petit os tégumentaire pousse jusqu'au marteau, formant son processus médian, on pense que ce dernier correspond à l'os articulaire tégumentaire (goniale) des reptiles. Situé à côté de ce dernier os, l'os angulaire tégumentaire des reptiles passe également dans la région de l'oreille, devenant l'os tympanique (tympanique). Ces changements sont visibles dans le schéma ci-joint (Fig. 541).
Les données paléontologiques confirment la possibilité de transformations cachées ci-dessus. Parmi les bêtes à dents animales (Theriodontia), il y a celles chez lesquelles la dentition est particulièrement développée, atteignant l'arrière de la jonction de la mâchoire inférieure avec une brosse carrée. Dans le même temps, l'os squameux descend le long de l'os carré jusqu'à la même articulation. Les os dentaires et squameux se rapprochent ainsi de plus en plus.
Enfin, dans certains os à dents animales, les os dentaires et squameux participent à l'articulation avec les os articulaires et carrés.
Quant à l'appareil hypoglosse des mammifères, il est constitué d'un corps, d'une partie basale modifiée (copule) de l'arc hyoïde, des cornes antérieures (cornua anteriora), formées par la partie inférieure de l'arc hyoïde, et des cornes postérieures ( cornua posteriora), transformé à partir du premier arc branchial. Les cornes antérieures sont constituées de plusieurs ossifications (jusqu'à quatre) ; le segment faisant face au crâne se développe souvent jusqu'à l'os pierreux, formant un processus styloïde (proc. styloideus).
Les deuxième et troisième arcs branchiaux ont donné lieu à la formation du cartilage thyroïde (cartilago thyreoidea.)
Ceintures de membres. La ceinture scapulaire des mammifères se caractérise principalement par la réduction des os du corbeau (procoracoideum, epicoracoideum, coracoideum), la tendance à la disparition des clavicules (clavicule) et la crête bien développée de l'omoplate (spina scapulae), se prolongeant dans la processus d'acromion (acromion).
Cependant, chez les monotrèmes, la ceinture scapulaire ressemble beaucoup aux reptiles. L'omoplate est équipée d'une protubérance acromiale, à laquelle la clavicule est adjacente à l'avant, et les os de corbeau larges et courts (coracoideum), allant au sternum, sont attachés à l'arrière de l'omoplate. Devant eux, il y a de larges procoracoïdes (procoracoideum) sous la forme d'os indépendants. Les clavicules (clavicule) sont reliées médialement à l'épiphore (épisternum) (Fig. 542). Chez d'autres mammifères, les cracoïdes sont pondus sous la forme de deux ossifications, mais ils perdent le contact avec le sternum et se développent jusqu'à l'omoplate, formant un processus coracoïde (proc. Coracoideus); les procoracoïdes disparaissent complètement (Fig. 543), les clavicules disparaissent chez certains prédateurs et ongulés en raison de l'adaptation à la course rapide, et chez les cétacés et les sirènes en raison de la transformation des membres antérieurs en nageoires.

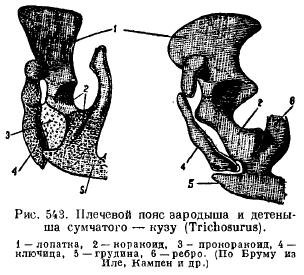
La ceinture pelvienne des mammifères présente une disposition typique des éléments lorsqu'elle est posée; sa caractéristique est, premièrement, la présence d'un os supplémentaire dans le cotyle (acetabulare), et deuxièmement, la fusion de tous ces os de chaque côté en un os sans nom (innominatum) et la fusion entre les os pubiens et ischiatiques, de sorte que un joint fermé est formé de tous les côtés du trou entre ces os (pour. obturatum) (Fig. 544). Chez les monotrèmes et les marsupiaux, un os spécial est attaché à l'os pubien par le bas pour soutenir le sac (marsupiale).
Squelette de membre dans les formes originales des mammifères, cela est dû à leur adaptation aux mouvements rapides - courir à l'aide de quatre membres, qui sont situés sous le corps, de sorte que ce dernier ne pende pas entre les membres, comme chez la plupart des reptiles, mais repose directement sur eux. La capacité de courir est due à la position différente des membres chez les mammifères. C'est avec eux que les membres sont orientés de sorte que les articulations du coude des membres antérieurs soient dirigées vers l'arrière et les articulations du genou des membres arrière soient dirigées vers l'avant. Lors du déplacement, les membres antérieurs sont projetés vers l'avant et tirent le corps en pliant le poignet et articulations du coude; les membres postérieurs poussent le torse vers l'avant en redressant la jambe au niveau de l'articulation du genou.
En termes de nombre d'os qui composent les membres et de leur emplacement, le squelette des membres des formes ancestrales de mammifères a une structure très typique d'un membre primitif à cinq doigts et a subi grands changements dans la classe que dans le processus d'adaptation à divers types de mouvements dans conditions différentes Mercredi.
Le membre antérieur est caractérisé par la présence dans la partie distale humérus trous (for.entepicondylare), la présence d'une protrusion proximale de l'ulna (olécrâne), la présence d'un os pisiforme (pisiforme) et la fusion des quatrième et cinquième os distaux du carpe (carpalia dis talia 4 + 6 = hamatum, ou uncinatum) et le centre proximal avec l'intermédiaire (centra dist. + intermedium = lunatum, ou semilunare). Du fait que la nomenclature des os du poignet des mammifères, adoptée en anatomie humaine, est souvent utilisée, nous donnons un tableau de ces noms dans le schéma indiquant l'emplacement des os.
Le membre postérieur des mammifères se caractérise principalement par le fait que la cuisse est tournée par l'articulation du genou devant la cuisse et présente des processus bien développés sur la partie proximale - trochanters, au nombre de 2-3 (trochanter majeur, mineur et tertius ); dans le bas de la jambe, le tibia est plus développé, il existe une rotule osseuse (patella), qui protège l'articulation du genou. Dans le tarse, il n'y a que deux os proximaux, bien développés : l'interne - le talus (talus = astragale), formé de la fusion des proximaux intermédiaire et central (intermedium + centrale proximale), et le calcanéum externe (calcanéus) , correspondant à l'os fibulaire du tarse (fibulare). À partir des os de la rangée médiane, le distal central (centrale distale) s'est transformé en scaphoïde (naviculare = scaphoideum). L'os tibial est soit réduit, soit rattaché à ce dernier.
La disposition des os du tarse et du métatarse et les synonymes des noms sont les suivants :
En relation avec l'adaptation au mouvement dans divers environnements, les membres des mammifères ont subi des changements très importants, et ces changements peuvent être particulièrement bien tracés sur le squelette. Malheureusement, nous ne disposons pas de données paléontologiques sur la structure des membres des ancêtres des mammifères. Il ne fait aucun doute que les premiers mammifères avaient des membres peu spécialisés du type décrit ci-dessus. Ce sont de tels membres que les marsupiaux les plus primitifs des mammifères modernes ont, par exemple, des opossums - des rats marsupiaux (Didelphys) et des insectivores menant un mode de vie arboricole. Cependant, il serait faux d'en conclure que les ancêtres des mammifères étaient certainement les habitants des forêts et, de plus, les formes qui grimpaient aux arbres. De plus, le membre de l'opossum, bien qu'il serve à se déplacer le long des branches des arbres, n'est en aucun cas un membre d'arbre spécialisé typique, mais conserve son type universel à cinq doigts. De tels membres pouvaient mieux survivre chez les animaux qui non seulement couraient sur le sol, mais aussi grimpaient aux arbres, ainsi que chez les animaux qui escaladaient les montagnes. La spécialisation des membres mammifères s'est déroulée simultanément avec la différenciation des ordres et d'autres unités systématiques plus petites, et la convergence a eu lieu.
Les membres subissent des modifications particulièrement profondes lorsqu'ils s'adaptent au vol, à la nage dans l'eau, aux déplacements souterrains ; les mouvements lents du sol et les modes de vie arboricoles ne sont pas associés à une spécialisation profonde.
Chez les chauves-souris (Chiroptères), le membre antérieur se transforme en aile, et la voilure est tendue entre quatre (2-b) doigts fortement allongés, dépourvus de griffes ; le premier orteil est court et a une griffe fortement développée à l'extrémité ; ce doigt ne joue aucun rôle dans le vol, il est utilisé pour l'escalade ; au poignet, les os scaphoïde, lunaire et central se confondent en un seul; l'avant-bras est allongé et se compose d'un radius bien développé, tandis que le cubitus est réduit et s'accumule au radius. Le profil aérodynamique engloutit également les pattes arrière. Entre ces derniers, il est différemment développé chez les chauves-souris (Fig. 545).
Chez les mammifères nageant dans l'eau, les membres se transforment en nageoires de la même manière que nous l'avons déjà vu chez les reptiles. Dans le même temps, les membres des baleines et des sirènes ont particulièrement changé. Contrairement à l'aile, le membre antérieur est considérablement raccourci dans sa partie proximale. Chez les baleines, les os de l'épaule et de l'avant-bras se sont transformés en plaques courtes et larges qui offrent de la place à une musculature puissante pour s'attacher. Toutes les parties du membre sont reliées les unes aux autres sans bouger, formant une forte pagaie. Dans ce cas, le nombre de phalanges peut être considérablement augmenté. Nous avons vu les mêmes changements chez les reptiles. L'augmentation du nombre de phalanges dépend du fait que la diaphyse des phalanges ne se développe pas avec les glandes pinéales et s'ossifie à partir de centres séparés. Une telle ossification séparée est due au processus retardé d'ossification chez ces animaux. La lenteur du processus d'ossification conduit au fait que la plupart des os à leur jonction restent cartilagineux.
Les nageoires des sirènes (Sirenia) ont une ressemblance extérieure avec celles des cétacés : l'épaule et l'avant-bras sont articulés entre eux, de sorte qu'ils peuvent se plier ; il y a presque tous les os du poignet et les cinq doigts avec un nombre légèrement modifié de phalanges (Fig. 546).
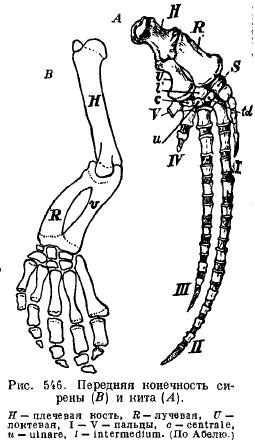
Membres postérieurs en baleines modernes et les sirènes sont réduites de sorte qu'il n'en reste que les restes du bassin (Fig. 547 et 548).
Un autre type d'adaptation des membres au mouvement dans le foyer est observé chez les pinnipèdes (Pinnipedia). Ici, la queue disparaît presque et les pattes postérieures reculent, jouant un rôle organe moteur... Les trois familles de cet ordre représentent, selon le degré de développement de l'adaptabilité au mouvement dans l'eau, trois stades pour ainsi dire (non génétiquement). Chez les morses et les phoques à oreilles (Otariidae), les membres postérieurs peuvent encore se plier vers l'avant sous le corps, mais chez les morses, la structure des membres est plus primitive, ils sont plus courts et plus mobiles. Chez les phoques (Phocidae), les pattes postérieures ne peuvent plus se plier en avant sous le corps, cependant, les griffes sont toujours conservées sur les orteils (nageoires).
Chez les animaux fouisseurs, les membres subissent également de très fortes modifications : les membres de la taupe marsupiale (Notoryctes typhlops) ont particulièrement changé. Il a deux arêtes sur son omoplate, comme beaucoup d'animaux fouisseurs, ce qui est dû au fort développement du muscle triceps de l'épaule ; a une clavicule, comme tous les mammifères fouisseurs. L'humérus a une crête très forte qui se ramifie en forme de fourche et sert à attacher un deltoïde fort (nécessaire pour creuser) et muscles pectoraux; le condyle interne de l'épaule est très développé en raison de fort développement les muscles fléchisseurs de l'ulna (proc. olecranon), qui sert de point d'attache pour le muscle triceps ; le poignet est singulièrement changé, devenant très fort ; Les 1er et 2e doigts sont repoussés du côté palmaire, et seuls les 3e et 4e, munis d'énormes griffes, surtout le 3e, servent à creuser ; le nombre de phalanges au niveau des doigts est très faible : 2 et 1. 5e métacarpien avec une large griffe plate, ce qui augmente la surface d'enfouissement ; dans le plan de la paume, il y a un os sésamoïde, auquel est attaché un muscle qui fléchit les doigts, qui joue un grand rôle dans le creusement; il augmente la force du poignet; comme d'autres formes fouisseuses, la symphyse os pubiens très court; l'ischion se développe avec les apophyses transverses des deux dernières vertèbres sacrées ; sur la cuisse, le genou et le tibia, les crêtes et les excroissances sont très développées - les lieux d'attache des muscles; le tibia et le péroné (tibia et péroné) sont fermement fusionnés, comme une taupe dorée (Chrysochloris); la semelle est repliée vers l'extérieur, comme chez Chrysochloris ; Le 5e orteil est replié sur la surface plantaire, le 5e ongle du pied est large et spatulé, le 4e est long et en forme de faucille, le 3e et le 2e deviennent plus courts et plus larges ; sur le sternum une crête très développée, comme la taupe dorée, le rat taupe (Spalax) et la taupe (Talpa) (Fig. 549).
Le fort développement de crêtes sur les os pour la fixation des muscles, le raccourcissement du carpe, la connexion ferme des carpiens, la réduction de certaines phalanges et le renforcement d'autres qui jouent un rôle dans le creusement - tous ces signes sont caractéristiques de mammifères fouisseurs en général. Mais selon le type de sol que l'animal creuse, il y aura des différences. En creusant dans un sol meuble (par exemple, dans une taupe - Talpa) (Fig. 550), la main se dilate fortement, elle agit comme une pelle et a tendance à développer le 6e orteil. Le membre antérieur d'une taupe est comparable à une pelle utilisée pour creuser dans un sol meuble. Au contraire, dans les formes qui creusent dans la terre ferme, la main devient plus étroite, seuls quelques doigts participent au creusement, principalement les 3e et 4e, se développant plus fort que d'autres, comme chez Notoryctes et Chrysochloris. Cette brosse est comparable au tampon à creuser pour sol dur.
![]()
On voit la même chose chez les mammifères déchiqueter les structures solides des termites, par exemple chez le fourmilier (Myrmecophagus), chez le tamandua (Tamandua). Ces animaux marchent en s'appuyant sur le bord extérieur des pattes, les griffes repliées.
L'adaptation des mammifères aux mouvements rapides sur le sol a provoqué des changements très importants dans les membres. Le mouvement au sol, caractéristique des mammifères primitifs, s'effectuait en s'appuyant sur l'ensemble de la main et du pied ; formes originales les mammifères étaient principalement plantigrades ou plantigrades, c'est-à-dire qu'ils s'appuyaient sur tous leurs pieds pour marcher, comme, par exemple, les ours. Mais une telle structure ne permet qu'une marche relativement lente par rapport aux antilopes aux pieds rapides ou aux chevaux, ou aux lièvres sauteurs, kangourous, gerboises. Au cours du développement phylogénétique, nous voyons une élévation progressive au-dessus du sol du poignet et du tarse (carpe et tarse), qui étaient ensuite suivis par le métacarpe et le métatarse, mais les phalanges touchaient toujours entièrement le sol. À la fin, les phalanges se sont également redressées, les membres antérieurs et postérieurs ont commencé à reposer sur le bout des doigts. On peut désormais retracer pas à pas les étapes de cette évolution non seulement chez les ongulés, mais aussi chez les mammifères carnivores qui ont acquis la capacité de courir vite. Au fur et à mesure que les membres se redressent, les doigts extrêmes cessent de toucher le sol, cessent de fonctionner et s'atrophient. Il en résulte une plus petite surface de contact avec le sol. Les modifications de la partie distale des membres ont entraîné des modifications de la partie proximale: les carpes et les tarsales sont de plus en plus proches l'une de l'autre, le cubitus et le péroné sont réduits, les membres ne commencent à se déplacer que dans le plan sagittal, les métapodes médians se fondent dans un os.
Deux voies de développement mènent à la formation des membres des ongulés modernes. Dans une rangée, l'axe du membre passe au milieu du troisième orteil et ce dernier se développe le plus fortement, tandis que le reste devient rudimentaire. Ce sont des équidés (Perissodactyla), mésaxiaux (Mesaxonia). Dans d'autres, l'axe passe entre le troisième et le quatrième doigt, se développant uniformément: artiodactyles (Artiodactyla), paraxial (Paraxonia). Les doigts latéraux sont réduits. Le développement des équidés est plus clairement visible dans le développement phylogénétique des membres du cheval (Fig. 551). Le développement de l'état des sabots fourchus était dû, entre autres, au fait qu'il était précédé de la réduction du premier orteil, de sorte que dans la transition de la marche à pied, il n'a presque pris aucune part, alors que chez les ancêtres des animaux à un sabot, le développement du cinquième orteil était initialement normal.
Il est intéressant de noter que l'artiodactyle et l'artiodactylaire ont émergé à plusieurs reprises au cours du processus évolutif et, dans certains cas, ont conduit à un type très spécialisé, par exemple chez les Proterotlieriidae d'Amérique du Sud (éteints).
Les orteils restants sont recouverts de sabots, qui peuvent se différencier davantage en fonction du mode de vie et de l'habitat. Par exemple, les chèvres qui s'accrochent aux rochers ont des sabots raides et étroits avec une arête vive ; à renne ils sont très longs, largement bifurqués et servent de sortes de skis qui empêchent l'animal de tomber dans la neige et les marécages.
Simultanément aux doigts, avec le développement de la marche sur les doigts, un changement se produit au niveau du poignet (carpe) et du tarse (tarse). Cela se passe différemment chez les artiodactyles et les équidés en raison d'une répartition différente de la gravité. Enfin, en lien avec le passage au nalcewalking, la valeur du cubitus (ulna) comme support de l'épaule diminue progressivement : elle diminue ou se confond avec le radius (radius). De même, le péroné est réduit et disparaît finalement dans les membres postérieurs.
La signification et la nature des changements décrits sont clairement visibles à partir de la comparaison des membres d'un chameau et d'un éléphant (Fig. 552). Chez un éléphant, la main (pied) reste courte, et l'épaule et l'avant-bras (cuisse et bas de la jambe) sont allongés. Au contraire, chez le chameau, les métapodes accrètes sont allongés ; le haut du bras et l'avant-bras (ainsi que la cuisse et le bas de la jambe) restent courts. En raison de la perte d'un certain nombre d'os, le squelette du membre est simplifié, ce dernier devient plus léger. La réduction des doigts entraîne moins de friction, le risque de blessure, de rupture des ligaments est réduit. La fusion des métapodes crée une grande force. Le membre de l'éléphant est beaucoup plus lourd. L'abondance des os dans la main nécessite la connexion de tous les doigts pour obtenir la stabilité. peau générale... Le membre de l'éléphant est une colonne d'une grande force, mais sans la légèreté du membre du chameau.
Une autre façon de se déplacer rapidement au sol est de sauter. Cette adaptation est apparue à plusieurs reprises dans l'histoire des mammifères et, d'ailleurs, dans une grande variété de troncs phylogénétiques : chez les marsupiaux - chez les kangourous (Macropus), chez les insectivores - chez les sauteurs (Macroscelidae), et, enfin, chez les différents rangs développement des rongeurs : ceci. les lièvres (Leporidae) de Duplicidentata ; dans la série des porcs-épics (Hystricomorplia), un groupe d'arpenteurs (Pedetinae) s'est développé; parmi les gerboises ressemblant à des souris (Myomorpha), nous avons la gerboise australienne (Conilurus), les gerboises de l'Ancien Monde (Jaculus, Alactaga), les sauteurs américains (Zapus) et les sauteurs en maille (Dipodomys).
Le développement de la capacité de sauter est associé à l'allongement du membre postérieur, à savoir les os du métatarse (métatarse). De plus, cela entraîne la perte de doigts extrêmes et les os du tarse sont très fermement connectés les uns aux autres. Les pattes du lièvre de terre (Alactaga) et les vraies gerboises (Dipus) en sont un exemple.
Les pattes antérieures d'Alactaga sont petites et courtes. Le lièvre de terre les utilise pour creuser, mais ne touche pas le sol en sautant. Le saut s'effectue uniquement à l'aide des pattes postérieures et même sans l'aide de la queue. Ce dernier ne touche le sol que dans une position calme, lorsque l'animal repose sur sa queue. Le squelette de la jambe a la particularité que les os métatarsiens des 2e, 3e et 4e orteils fusionnent en un seul os. Les extrémités des os métatarsiens ci-dessous ne sont pas fusionnées et la médiane (3e orteil) fait saillie plus en avant. Des phalanges qui leur sont reliées, celles du milieu sont encore les plus longues. Les 1er et 5e orteils sont rudimentaires et ne touchent pas le sol. Ces deux doigts ont longueur égale en raison du fait que l'os métatarsien pouce plus long que le 5. Le premier en a 2, le second - 3 phalanges.
A Dipus, la spécialisation est allée encore plus loin : le 1er doigt a disparu sans laisser de trace, du 5e il ne reste qu'un rudiment. Majeur deux fois plus faibles que les deux extrêmes, et à peine plus longs qu'eux. Les orteils sont parallèles. Le majeur est réduit car sa fonction est prise en charge par le 2ème et le 4ème, aux extrémités desquels il y a un accent lors de l'assise et du saut. Dans une évolution ultérieure, le 3e orteil devrait disparaître : nous avons un type d'adaptation inachevé chez Dipus.