Ինչ է կաթնասունների կրծքային ողնաշարը: Կաթնասունների արտաքին կառուցվածքը, կմախքը և մկանները ընտանի շան օրինակով
Ողնաշարը բաղկացած է արգանդի վզիկի, կրծքային, գոտկային, սակրալ և պոչային շրջաններից։ Նրան բնորոշ հատկանիշ- ողերի պլատիցելիային (հարթ մակերևույթներով) ձև, որոնց միջև կան աճառներ. միջողային սկավառակներ... Վերին կամարները լավ արտահայտված են: Արգանդի վզիկի շրջանում կան յոթ ողեր, որոնց երկարությունը կախված է պարանոցի երկարությունից; միայն ծովախորշում և ծույլում՝ Choloepus hof-fmani-ում, դրանք 6-ն են, իսկ ծույլում՝ Բրադիպուսը՝ 8-10: Արգանդի վզիկի ողերը շատ երկար են ընձուղտում և շատ կարճ կետաձևերի մոտ, որոնք չունեն արգանդի վզիկի հատված: Կրծքավանդակը կազմող կողիկներն ամրացված են կրծքային շրջանի ողերին։ Այն փակող կրծքավանդակը հարթ է և միայն մոտ չղջիկներև հզոր առջևի վերջույթներ ունեցող փորված տեսակները (օրինակ՝ խալերը) ունեն փոքրիկ գագաթ (քիլ), որը ծառայում է որպես կրծքավանդակի մկանների կցման կետ: Կրծքավանդակի շրջանում կան 9-24 (ավելի հաճախ՝ 12-15) ողեր, վերջին 2-5 կրծքային ողերը կրում են՝ կեղծ կողոսկրեր՝ կրծոսկրին չհասնող, գոտկային շրջանում՝ 2-ից 9 ողեր; տարրական կողոսկրերը միաձուլվում են իրենց լայնակի մեծ պրոցեսների հետ։ Սակրալ շրջանը ձևավորվում է 4-10 միաձուլված ողերով, որոնցից միայն առաջին երկուսն են իսկապես սակրալ, իսկ մնացածները՝ պոչային։ Ազատ պոչային ողերի թիվը երկարապոչ մողեսում տատանվում է 3-ից (գիբոնում) մինչև 49:
Առանձին ողերի շարժունակության աստիճանը տարբեր է։ Փոքր վազող և մագլցող կենդանիների մոտ այն մեծ է ողնաշարի ամբողջ երկարությամբ, ուստի նրանց մարմինը կարող է թեքվել տարբեր ուղղություններև նույնիսկ գլորվել գնդակի մեջ: Կրծքավանդակի և գոտկային շրջանների ողնաշարերն ավելի քիչ շարժուն են խոշոր, արագ շարժվող կենդանիների մոտ։ Կաթնասունների մեջ, որոնք ճանապարհորդում են հետևի ոտքերը ah (կենգուրուներ, ջերբոաներ, ցատկերներ), ամենամեծ ողերը գտնվում են պոչի և սրբանի հիմքում, և հետագայում դրանց չափը աստիճանաբար նվազում է: Սմբակավոր կենդանիների մոտ, ընդհակառակը, ողնաշարերը և հատկապես նրանց ողնաշարային պրոցեսներն ավելի մեծ են կրծքային շրջանի առաջի մասում, որտեղ նրանց կցված են պարանոցի և մասամբ առաջնային վերջույթների հզոր մկանները (
Շան արտաքին կառուցվածքը
Շունն առաջին ընտանի կենդանին էր։ Մարդը նրան ընտելացրել է հին ժամանակներում: Շունը որսի ժամանակ օգնել է պարզունակ մարդուն, հսկել նրա տունը։ Այժմ հայտնի են ծառայողական շներ, որսորդական և դեկորատիվ, ինչպես նաև խառնաշփոթներ (խառնաշփոթ): Մեր Հայրենիքի սահմանները պահպանելու ծառայությունն իրականացնելու հարցում օգնում են շները, և Մեծի ժամանակ Հայրենական պատերազմորոնել է վիրավորներին, օգնել հետախուզության և կապի մեջ. Շների վրա գիտնականները ուսումնասիրություններ են անցկացնում մարմնի կենսագործունեության վերաբերյալ: Շուն մարդու առաջայցելած տարածք.
Շան մարմինը, ինչպիսին է հովիվը կամ խոզուկը, սլացիկ է և մկանուտ։ Նրա վերջույթները գտնվում են ոչ թե մարմնի կողքերում, ինչպես սողունների մոտ, այլ մարմնի տակ, ուստի կենդանու մարմինը չի դիպչում գետնին։ Շունը քայլում է ամուր ճանկերով հենվելով մատների վրա։ Ճկուն պարանոցը նպաստում է գլխի մեծ շարժունակությանը: Կենդանու բերանը սահմանափակված է շարժական շրթունքներով՝ վերին և ստորին: Վերևում վերին շրթունքկա քիթ՝ զույգ արտաքին քթի բացվածքով՝ քթանցքերով։ Աչքերը լավ զարգացած կոպեր ունեն։ Թարթող թաղանթը (երրորդ կոպերը) շան մեջ, ինչպես բոլոր կաթնասունների մոտ, թերզարգացած է։ Բոլոր կենդանիներից միայն կաթնասուններն ունեն արտաքին ականջ - Աուրիկուլ... Շան արտաքին ականջը մեծ է և շարժական։
Կաթնասունների ծածկոցներ
Կաթնասունների մաշկը ամուր է և առաձգական, կենդանիների մեծ մասի մեջ դրված են մազերի հիմքերը, որոնք կազմում են ողնաշարավորների այս դասի բնութագիրը։ մազերի գիծ ... Տարբերակել հաստ ու երկար մազեր — ողնաշարըև ավելի կարճ, ավելի մեղմ - հատակին, կամ ներքնազգեստ... Կոպիտ և դիմացկուն ծածկը պաշտպանում է ներքնազգեստը և մաշկը վնասվելուց: Ներքնազգեստը, որը շատ օդ է պահում, լավ է պահպանում մարմնի ջերմությունը։ Կենդանիները, բացի մորթուց և մորթուց, զարգանում են մեծ մազեր- շոշափման օրգաններ.
Կաթնասունների մազերը, ինչպես թռչունների փետուրները և սողունների թեփուկները, կազմված են եղջյուրավոր նյութերից։ Տարին երկու անգամ շունը, ինչպես շատ այլ կաթնասուններ, թափվում է. նրա մազերի մի մասը թափվում է և փոխարինվում նորերով: Մազերի հիմքը գտնվում է հատուկ մազի պարկի ներսում, որտեղ բացվում են մոտակա ծորանները ճարպագեղձեր... Նրանց սեկրեցները յուղում են մաշկը և մազերը, ինչը նրանց դարձնում է առաձգական և չթրջված ջրով:
Կաղապար. Կաթնասունի մաշկի և մազերի կառուցվածքը
Կենդանիների մեծ մասի մաշկի մեջ գտնվում են և քրտնագեղձեր ... Քրտինքը գոլորշիանում է մարմնի մակերեսից և սառեցնում այն։ Քրտինքի հետ միասին օրգանիզմից արտազատվում են նաև ավելորդ աղը և միզանյութը. Այսպիսով, քրտինքի խցուկները դեր են խաղում լրացուցիչ մարմիններարտանետում.
Շունը քրտնագեղձերքիչ է, և մարմնի սառեցումը ձեռք է բերվում շնչառության ավելացումով:
Կաթնասունների մատների ծայրերում կան եղջյուրավոր ճանկեր, եղունգներ կամ սմբակներ։ Երբեմն եղջյուրավոր գոյացություններ են առաջանում նաև գլխի վրա (եղջյուրներ ռնգեղջյուրների, անտիլոպների, խոշոր եղջերավոր անասունների և այլն) կամ պոչի վրա (օրինակ՝ առնետների մոտ՝ բեղջավոր թեփուկները)։
Կաթնասունների կմախք
Կաթնասունների կմախքը բաղկացած է նույն հատվածներից, ինչ մյուս ողնաշարավորների մոտ։ Կենդանիների գանգն առանձնանում է ավելի մեծ գանգուղեղով, որը կապված է մեծ չափսուղեղը. Կաթնասունների համար շատ բնորոշ է արգանդի վզիկի 7 ողերի առկայությունը։ Ե՛վ երկար պարանոցով ընձուղտները, և՛ կետերն ունեն նույն թվով արգանդի վզիկի ողերը։ Կրծքային ողերը (սովորաբար 12-15) կողերի և կրծքավանդակի հետ միասին կազմում են ամուր կուրծք։ Զանգվածային ողնաշարեր գոտկայինշարժականորեն հոդակապված միմյանց հետ: Այս հատվածում մարմինը կարող է թեքվել և թեքվել: Գոտկատեղի ողերի թիվը տարբեր է տարբեր տեսակներ(2-9), շունն ունի 6: Սակրալ ողնաշարը (3-4 ողնաշար) միաձուլվում է կոնքի ոսկորների հետ: Պոչի ողերի քանակը (երեքից մինչև մի քանի տասնյակ) կախված է պոչի երկարությունից։

Նկար. Կաթնասունների կմախքը՝ օգտագործելով շան օրինակը

Կաղապար՝ շան գանգ
Առաջնային գոտիԿաթնասունները բաղկացած են երկու ուսի շեղբերից, որոնց վրա կցված են ագռավի ոսկորները և երկու մանյակ: Շան ոսկորները զարգացած չեն։ Հետևի վերջույթների գոտի- pelvis - ձևավորվում է երեք զույգով կոնքի ոսկորներ... Կաթնասունների և սողունների տարբեր տեսակների վերջույթների կմախքները նման են, սակայն տարբեր տեսակների մոտ դրանց կառուցվածքի մանրամասները տարբեր են և կախված են կենդանու կենսապայմաններից։
Կաթնասունների մկանները
Կաթնասունների մեծ մասը, ներառյալ շունը, հատկապես զարգացած են մեջքի, վերջույթների և նրանց գոտիների մկանները: Շունը կարող է վազել մեծ ցատկերով՝ թեքելով և ուղղելով մարմինը, հերթափոխով գետնից հրելով իր առջևի կամ հետևի ոտքերով: Ուժեղ մկանները շարժում են ստորին ծնոտը: Ատամներով զինված ծնոտները սեղմելով՝ շունը ամուր բռնում է որսին։

Կաղապար: Շան մկանները շարժում են ստորին ծնոտը
Ուսի գոտի կաթնասունները պարզեցված է, բաղկացած է մեծ ուսի շեղբերունենալով կորակոիդային աճ, և կլավիկուլ(նկ. 80, 81): Զարգացման սաղմնային փուլում և հասուն մոնոտրեմներում կորակոիդանկախ ոսկոր է:
Scapulaգտնվում է վերևում կրծքավանդակը, ունի եռանկյունաձև ձև և բարձր ելք (գագաթ): ակրոմիալմկանների ամրացման գործընթաց: Սկապուլայի կորակոիդային աճի և ակրոմիալ պրոցեսի տեղակայման վայրում կա glenoid fossaառջեւի վերջույթը ամրացնելու համար.
Կոլարոսկրձողաձևը ծառայում է ուսի և կրծքավանդակի միացմանը: Կլավիկուլն առավել զարգացած է առջևի վերջույթներ կատարող կենդանիների մոտ: բարդ շարժումներ(չղջիկներ, խալեր, պրիմատներ, կրծողներ և այլն): Շատ տեսակների մոտ, որոնք բնութագրվում են միապաղաղ շարժումներով (սմբակավոր կենդանիներ, շներ) բացակայում է վզնոցը։ Ընդհանուր առմամբ, առջեւի վերջույթների գոտին մկանների եւ կապանների միջոցով կապված է առանցքային կմախքի հետ։
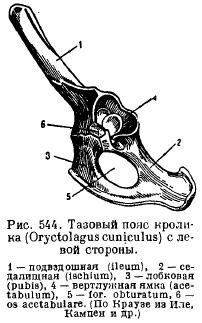
Կոնքի գոտի (նկ. 82 Ա) կազմված է զույգ անանուն ոսկորներհոդակապված iliac, sciatic եւ pubicոսկորներ. կոնք փակ տեսակիև իլիումի միջոցով ամուր կապված է սակրալողնաշարը. Երբ կոնքի տարրերը միասին աճում են, pubic միությունև ձևավորվեց acetabulumհետևի վերջույթների ամրացման համար. Օղակաձեւ ոսկորի ստորին հատվածում կա խցանման անցք.
Զուգակցված վերջույթների կմախք
Կաթնասունների վերջույթները, ինչպես ցամաքային այլ ողնաշարավորների անդամները, հնգմատանի են և ներկայացնում են եռանդամ լծակ, որի բոլոր մասերը շարժական կերպով կապված են միմյանց հետ։
Առջևի վերջույթ ներառում է ուս, նախաբազուկ և ձեռք(նկ. 81): Brachial ոսկորլավ զարգացած, ունի կլորացված գլուխ՝ վերջույթի գոտին ամրացնելու համար։ Նախաբազուկկազմել երկու ոսկոր - ճառագայթային եւ ulnarունենալով օլեկրանոնվերեւում. Ուսի և նախաբազկի միացման տեղում է արմունկի համատեղ:
Խոզանակ ներառում է երեք բաժին՝ դաստակ, մետակարպուս և ֆալանգներ. Դաստակբաղկացած է երեք շարքով դասավորված 8-10 տարրերից ... Pasternներառում է հինգ ոսկոր (ըստ մատների քանակի): Յուրաքանչյուր մատն ունի երեք ֆալանգ, բացառությամբ առաջին մատի, որն ունի երկու ֆալանգ: Ի տարբերություն սողունների և թռչունների միջկարպային հոդերի, շարժ համատեղտեղակայված կաթնասունների մեջ նախաբազկի և ձեռքի միջև(ինչպես երկկենցաղները):
Հետևի վերջույթ Կաթնասունները նույնպես ունեն երեք բաժին. ազդր, ստորին ոտք և ոտք(նկ. 82 Բ): Ֆեմուրզանգվածային և կլորացված գլխով մտցվում է կոնքի գոտու ացետաբուլում: Սունկը ներառում է երկու ոսկոր. tibia եւ tibia.Ավելի մեծ սրունքը սրունքի ներքին տարրն է, իսկ թմբիկը նրա արտաքին մասն է:
Ազդրի և ստորին ոտքի միջև կա համատեղծածկված ծնկի գլխարկորոնք առաջացել են նրանց ոսկրացած մկանային ջլերից: Հոդային ոսկորների մակերեսները ծածկված են աճառային գոյացություններով։ Կոճ համատեղկաթնասունները գտնվում են սրունքի ոսկորների ստորին ծայրերի և ոտնաթաթի մոտակա հատվածի միջև։

Բրինձ. 82. Աղվեսի կոնքի գոտի (A) և հետևի վերջույթ (B).
1 – իլիում, 2 - ischium, 3 - pubic ոսկոր, 4 - acetabulum, 5 - femur, 6 - patella, 7 - tibia, 8 - fibula, 9 - tarsus, 10 - metatarsus, 11 - phalanges մատների, 12 - talus, 13 - calcaneus.
Տարզային տարրերոտքերը դասավորված են երեք շարքով և ձևավորում են աճ. calcaneusբնորոշ կաթնասուններին. Metatarsusբաղկացած է հինգ ոսկորներից, որոնց կցված են մատների ֆալանգները։ Ինչպես առջևի վերջույթներում, այնպես էլ յուրաքանչյուր մատի մեջ դրանք երեքն են, բացառությամբ առաջինի (ներքին), որն ունի ընդամենը երկու ֆալանգ։
Կաթնասունների համար բնորոշ է վերջույթների հատուկ դիրքավորումը մարմնի նկատմամբ՝ դրանք գտնվում են մարմնի տակ, ինչը նպաստում է շարժումների ավելի մեծ մանևրելուն։ Այս դեպքում արմունկի հոդը ուղղված է հետին, և ծնկահոդընդհակառակը, այն ուղղված է առաջից։
Կաթնասունների տեսակների բազմազանության, նրանց ապրելակերպի և շարժման եղանակների, վերջույթների հարաբերական երկարության, բաղկացուցիչ տարրերի հարաբերակցության, ոսկորների կազմաձևման, մատների քանակի և այլնի հետ էապես տարբերվում են: Օրինակ. չղջիկների մոտ՝ առաջնային վերջույթի երկարությունը, հատկապես մատների ֆալանգները, որոնք ծառայում են թռչող թաղանթի ամրացմանը։
Ընդհակառակը, խլուրդի առջեւի վերջույթը կրճատված է, ձեռքի ափի հատվածը՝ լայնացած, վեցերորդ մատը լավ զարգացած է։ Կետերի մոտ ձեռքը կտրուկ երկարացվում է մատների ֆալանգների քանակի ավելացման պատճառով, ինչի արդյունքում ձեռք բերված վերջույթը. մեծ նմանությունխաչաձուկ ձկան լողակով։ Մեծ կենսազանգվածով և արագ վազքով կենդանիների մատները (էկվիդներ, արտիոդակտիլներ) զգալիորեն կրճատվել և փոփոխվել են։
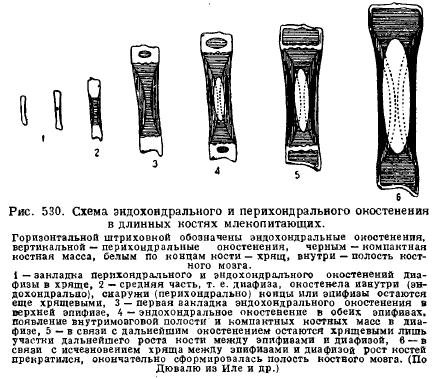
Կաթնասունների կմախքի առանձնահատկություններից է մեծ զարգացումաճառը երիտասարդ կենդանիների մեջ և դրա երկարաժամկետ պահպանումը զարգացման հետսեմբրիոնային շրջանում: Դա պայմանավորված է աճառի և ոսկորների աճի առանձնահատկություններով՝ կապված հենց կենդանու աճի հետ։ Հենց աճառն է աճում իր ողջ զանգվածով (ինտերստիցիալ), աճը տեղի է ունենում արագ, և մասերի համամասնությունները կարող են պահպանվել, մինչդեռ ոսկորն ավելի դանդաղ է աճում և միայն մակերեսից, այնպես որ կմախքի այն մասերը, որոնք միշտ հայտնվում են: մնալ նույն հեռավորության վրա, և ոսկորների աճի հետ մասերի համամասնության պահպանումն անհնար է դառնում։ Այդ իսկ պատճառով երիտասարդ կենդանիները միշտ պարունակում են մեծ քանակությամբ աճառ և հատկապես այն վայրերում, որտեղ աճը ուժեղ է։ Այսպիսով, երկար ոսկորներվերջույթները միշտ բաղկացած են միջին մասից՝ դիաֆիզից, որն առաջին հերթին ոսկրանում է, և ծայրերից, որոնք երկար ժամանակ պահպանվում են աճառով՝ սոճու գեղձից։ Դիաֆիզի ոսկրացումը սկսվում է առաջին հերթին ծայրամասից՝ պերիխոնդրալից, այսինքն՝ բջիջների շնորհիվ, որոնք ծագում են աճառը ծածկող կապի հյուսվածքից՝ պերիխոնդրիումից; ապա դրան միանում է աճառի ներքին ոսկրացումը՝ էնդոխոնդրալ ոսկրացում; միայն ավելի ուշ, երբ ոսկրածուծի խոռոչները հայտնվում են դիաֆիզում ոսկորների քայքայմամբ, սոճու գեղձերը սկսում են էնդոխոնդրիկորեն ոսկրանալ. դիաֆիզների և էպիֆիզների միջև, սակայն աճառը երկար է մնում, որի օգնությամբ ոսկորը երկարում է համամասնությունների պահպանմամբ (նկ. 530):
ՈղնաշարԿաթնասունները, ի տարբերություն Sauropsida-ի, բնութագրվում են ողերի միջև ազատ հոդերի բացակայությամբ, բացառությամբ առաջին և երկրորդ ողերի։ Ողնաշարային մարմինների միջև ընկած են միջողային առաձգական կապ հյուսվածքի սկավառակները: Ընդհանուր առմամբ, դա հանգեցնում է ողնաշարի ավելի քիչ ճկունության, ինչը պայմանավորված է վերջույթների օգնությամբ կաթնասունների ավելի մասնագիտացված տեղաշարժով:
Ողնաշարային մարմինների ոսկրացումը տեղի է ունենում աճառային սոճու գեղձի միջոցով: Մարմիններն ունեն կամ հարթ մակերեսներ, կամ թեթևակի ուռուցիկ, ժամանակակից սմբակավոր կենդանիների մեծ մասում դրանք թեթևակի օպիստոկելային են: Ողնաշարերի վրա լավ զարգացած են, հատկապես կրծքավանդակի շրջանում, ողնաշարային պրոցեսներ, ինչպես նաև լայնակի պրոցեսներ, որոնք նստում են աղեղների վրա. վերջինիս հիմքում առաջանում են հոդային պրոցեսներ։

Արգանդի վզիկի ողերի թիվը սովորաբար 7 է (նկ. 531)՝ անկախ պարանոցի երկարությունից, ուստի պարանոցի երկարությունը, ի տարբերություն թռչունների, կախված է բացառապես ողերի երկարությունից։ Միայն ծովախորշը (Manatus) և ծույլերի մեկ տեսակ (Choloepus hoffmani) ունեն 6 արգանդի վզիկի ողեր, իսկ եռոտանիները (Bradypus) ունեն 9։ , մասամբ միաձուլելով ողերը, մինչդեռ ընձուղտի պարանոցում այս 7 ողերը շատ երկար են։ Առաջին 2 արգանդի վզիկի ողերը ձևափոխվում են ատլասի և էպիստրոֆիայի՝ կապված իրական հոդերի հետ, իսկ մոնոտրեմներում օդոնտոիդ պրոցեսը դեռ գործում է հատուկ ոսկորի տեսքով և չի միաձուլվել, ինչպես մյուս կաթնասունների մոտ, մարմնի հետ։ էպիստրոֆիան։ Ողնաշարերի հետ միասին աճում են արգանդի վզիկի շրջանի տարրական կողիկներ, որոնց արդյունքում առաջանում են ծակոտկեն լայնակի պրոցեսներ։ Արգանդի վզիկի ողերի մասնակի կամ ամբողջական միաձուլումը տեղի է ունենում կետաձկան մեջ. jerboas- ում բոլոր արգանդի վզիկի ողերը աճում են միասին, բացառությամբ 7-ի, մարսուալ խալում (Notoryctes) դրանք միաձուլվում են 2-րդից 6-րդ, խալերում `3-ից 4-րդ:
Կրծքավանդակի ողերի թիվը, որոնք բնութագրվում են դրանց վրա կողոսկրերի կցվածությամբ, տատանվում է 9-ից [մեկ կետաձկանում (Hyperoodon) և armadillo-ում (Tatusia)] մինչև 25 [ծուլության մեջ], բայց սովորաբար դրանք լինում են 12-13: Կրծքային ողերի վրա պրոցեսները լավ զարգացած են՝ ողնաշարավոր (proc. Spinosus), լայնակի կամ դիապոֆիզներ (proc. Transversus), հոդային կամ zygapophyses (proc. Articularis); կողիկներն իրենց գլխով միացված են մեկ կամ երկու հարակից ողերի մարմնին, իսկ տուբերկուլյոզներով՝ լայնակի պրոցեսներին։ Դրանք բաղկացած են երկու մասից՝ ոսկրային կողոսկր՝ պրոքսիմալ և աճառային՝ հեռավոր։ Կրծքավանդակին հասնում են միայն առաջի կրծքային ողերի կողերը, հետին կողերն ազատ վերջանում են (կեղծ կողիկներ)։ Գոտկատեղի ողերը, որոնք բնութագրվում են լայնակի պրոցեսների բաղադրության մեջ այստեղ ընդգրկված կողերի բացակայությամբ, կաթնասունների մոտ 2-ից 9, սովորաբար 6-7:
Կաթնասունների սրբանային ողերը այն ողնաշարերն են, որոնք, միասին մեծանալով, կապվում են իլիումը... Կան 1-10 այդպիսի ողեր, բայց դրանցից միայն 1-2-ն են իսկական սրբանային ողեր, այսինքն՝ նրանք, որոնցում կոնքի հետ հոդակապումը տեղի է ունենում սրբանային կողերին համապատասխանող անկախ ոսկրացումների միջոցով. մնացած սրբանային ողերը կեղծ են, այսինքն՝ պոչային ողերը կպած սրբանին։
Պոչուկի շրջանում ընդգրկված ողերի թիվը չափազանց տարբեր է՝ 3-ից ( չղջիկները, գիբոն) մինչև 49 [մողես (Manis macrura)]:
Կրծքավանդակը(կրծքավանդակը) կաթնասունների զարգանում է կողոսկրերի փորային ծայրերի միաձուլումից, որոնք այնուհետեւ կազմում են մեկ չզույգված թիթեղ՝ բաղկացած մի շարք բաժանմունքներից (նկ. 532):
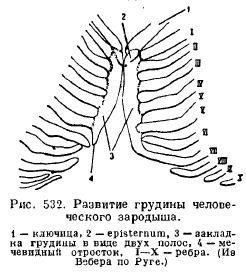
Միացված է կլավիկուլին (եթե այդպիսիք կան) և առաջին զույգ կողոսկրերի հետ երկարաձգված առաջային կրծքավանդակը կոչվում է բռնակ (praesternum, manubrium); որին հաջորդում է մարմինը կոչվող բաժանմունքը (mesosternum, corpus), որը հաճախ բաժանվում է մի շարք ոսկորների, որոնք հաջորդաբար տեղակայված են հետևյալ զույգ կողերի միջև. նույնիսկ ավելի հեռու ընկած է այսպես կոչված xiphoid գործընթացը (xiphistenum): Ոչ բոլոր կողիկներն են կապված կրծոսկրի հետ, քիչ թե շատ զգալի թվով կեղծ կողիկներն ազատ են վերջանում (նկ. 533):

Կլոակային կաթնասունների մոտ (Monotremata), կորակոիդների առկայության պատճառով, որոնք բացակայում են մնացած բոլոր կաթնասունների մոտ, կրծոսկրի բռնակի դիմաց դրվում է նախակրծունքի աճառը (prosternum), որը կապված է coracoids, որն այնուհետև տեղաշարժվում է զարգացող ոսկորով` էպիստերնումով (episternmn), որն ավելի է մոտեցնում կլոակային աճառը սողուններին:
ԳանգԿաթնասունները մի շարք հատկանիշներով տարբերվում են սողունների գանգից։ Սակայն այդ հատկանիշները ի հայտ եկան ոչ թե անմիջապես, այլ աստիճանական փոփոխության միջոցով, որը կարելի է հանդիպել՝ ուսումնասիրելով բրածո մողեսների (Theromorpha) գանգերը, հատկապես կենդանատամ կենդանիների կարգից (Theriodontia): Կաթնասունի գանգ - տրոպիբազային գանգ, բայց կարճացած միջուղղային մասով, որի պատճառով այն նման է պլատիբազալային, մեծ գանգուղեղով, առանձին ոսկորների միմյանց միաձուլման ընդգծված միտումով,
Կաթնասունների գանգի ոսկորների գտնվելու վայրը կարելի է դատել կից գծապատկերով (նկ. 534):
Կողմնային օքսիպիտալ ոսկորների վրա, հոդային պալարներից դուրս, սովորաբար տեղի է ունենում օքսիպիտալ պրոցես (pp. Paroccipitalis), որը հատկապես լավ զարգացած է կրծողների և շատ սմբակավոր կենդանիների մոտ։ Հոդային պալարների դիմաց սովորաբար բաց է լինում դեպի հիոիդ (XII) նյարդի արտաքին կողմը (for. Condyloideum anterius):
Լսողական պարկուճի աճառում դրված են մի քանի ոսկրացում, որոնք, սակայն, շատ վաղ միաձուլվում են երկու ոսկորների՝ առջևում պառկած՝ քարքարոտ (պետրոսում) և դրսում և հետևում՝ մաստոիդ (mastoideum); Այս երկու ոսկորները, միաձուլված միասին, տալիս են petro-mastoid կամ պարզապես քարքարոտ ոսկոր (petromastoideum, petrosum կամ perioticum), որը պարփակում է ներքին լսողական լաբիրինթոսը: Perioticum-ը հետևի մասում սահմանակցում է հիմնական և կողային օքսիպիտալ ոսկորների հետ, իսկ առաջի մասում՝ սֆենոիդ և թեւ-սֆենոիդ ոսկորներով; քանի որ անոթներն ու նյարդերն անցնում են այս ոսկորների և քարքարոտ ոսկորների միջև, այստեղ մնում են զգալի բացեր. հետևում - հետևի պատռված բացվածքը (lacerum posterius-ի համար), կամ պարանոցային բացվածքը (jugulare-ի համար) գլոսոֆարինգային (IX) նյարդի անցման համար, ճյուղեր: թափառող (X) նյարդի և օժանդակ (XI) նյարդի, ինչպես նաև ներքին պարանոցային երակի համար, իսկ առջևում՝ առաջի պատռվածքի անցքը (for. Lacerum anterius), որով ներքին քնային զարկերակը անցնում է գանգ։ . Երբեմն նավթային ոսկորի կապը գանգի հետ այնքան թույլ է, որ այն հեշտությամբ կարելի է առանձնացնել նրան կպած թմբկավոր ոսկորով, կամ նույնիսկ երբ գանգը թրծվում է, այն ամբողջությամբ թափվում է, ինչպես դա տեղի է ունենում կետաձկանների դեպքում։
Ականջի շրջանից դուրս ձևավորվում են երկու ծածկված ոսկորներ։ Դրանցից մեկը, որի ծագումը դեռ բավականաչափ պարզված չէ, զարգանում է արտաքին լսողական անցուղու տարածքում՝ կիսաշրջանաձև փոքր ոսկորի տեսքով։ Սա թմբկավոր ոսկորն է (tympanicum), որը, հավանաբար, անկյունային ոսկորի (angulare) նույնաբանն է: ստորին ծնոտըսողուններ, որոնք ծնոտի հոդակապման փոփոխությունների պատճառով ստորին ծնոտի այլ ոսկորների հետ տեղափոխվել են պարոտիդային շրջան: Գտնվելով լսողական անցուղու տարածքում ի սկզբանե կիսաշրջանաձև կամ օղակաձև ոսկորի տեսքով, այն հաճախ աճում և ձևավորում է, սովորաբար քարոտ ոսկորի հետ միասին, իսկ երբեմն թեփուկով, արտաքին լսողական միս (meatus auditorius externus): Թմբկաթաղանթը ձգվում է թմբկաթաղանթի վրա։ Ավելի խորը դեպի ներս, թմբկավոր ոսկորը նույնպես կարող է ներգրավված լինել միջին ականջի պատերի ձևավորման մեջ: Հաճախ լսողական շրջանի ստորին մասի թմբկավոր ոսկորը ուռչում է բարակ պատերով փուչիկի (bulla ossea) տեսքով:
Մաշկային ծագման երկրորդ ոսկորը, որը մասնակցում է ականջի շրջանի կմախքի ձևավորմանը, սակրավոր ոսկորն է (squamosum); այն բացառիկ զարգացում և նշանակություն է ստացել կաթնասունների մոտ՝ կապված նրան ստորին ծնոտի ամրացման հետ։ Կապիկների և մարդկանց մոտ ականջի շրջանի այս բոլոր ոսկորները միասին աճում են՝ կազմելով բարդ ժամանակավոր. թեփուկ ոսկորն այստեղ կազմում է միայն մի մասը, այն է՝ թեփուկները (squama temporalis) ժամանակավոր ոսկոր, մինչդեռ թմբկավոր ոսկորը թմբկավոր մասն է (pars tympanicum), իսկ պարոտիդ աճառային ոսկորներն իրենք են կազմում քարքարոտ (pars petrosa) և մաստոիդ (pars mastoidea) մասերը։ Շերտավոր ոսկորի զարգացման աստիճանը և նրա մասնակցությունը ուղեղային արկղի և լսողական մսի պատերի ձևավորմանը շատ տարբեր են, բայց ոսկորը միշտ ունի հոդային մակերես (savitas glenoidea) ստորին ծնոտի հոդակապման համար: Այսպիսով, կաթնասունների մոտ, ի տարբերություն մյուս բոլոր ողնաշարավորների, գոյություն ունի ստորին ծնոտի մի շատ հատուկ օրիգինալ կցորդ՝ ոչ թե քառակուսի ոսկորին, այլ գանգի շերտավոր ոսկորին։ Ստորին ծնոտի հոդային մակերեսը ընկած է թեփուկներից դեպի դուրս և առաջ ձգվող հիմքի վրա zygomatic գործընթաց(proc. zygomaticus): Հոդային մակերևույթից բարձր մոնոտրեմներում այն թափանցում է ժամանակավոր ջրանցքը (can.temporalis), որը հատկապես արտահայտված է պլատիպուսում։ Հոդային ֆոսայի հետևում կա պրոցես (proc. Postglenoideus):
Լսողական մասի դիմաց գանգի հիմքը ձևավորվում է հիմնական սֆենոիդ ոսկորով (basisphenoidale), վրա. ներքին մակերեսըորն ունի քիչ թե շատ զգալի դեպրեսիա՝ այսպես կոչված թուրքական թամբ (sella turtica), որի մեջ գտնվում է հիպոֆիզը։ Հիմնական սֆենոիդ ոսկորի կողմերում ընկած են՝ ձևավորելով գանգուղեղի առաջի պատի ստորին հատվածները, թև-սֆենոիդ ոսկորները (alisphenoidea), որոնք սովորաբար միաձուլվում են դրա հետ մեկ ոսկորի մեջ՝ ձևավորելով այս ոսկորի մեծ թևեր (alae magnae): Թև-սֆենոիդ ոսկորները հագեցած են երկու անցքերով՝ օվալ (for. Ovale)՝ եռաժանի նյարդի երրորդ ճյուղի (V3) անցման համար և կլոր (for. Rotundum)՝ նույն նյարդի երկրորդ ճյուղի անցման համար։ (V2). Այնուամենայնիվ, առաջինը հաճախ միաձուլվում է առջևի ծալքավոր բացվածքի հետ (for. Lacerum anteris), իսկ երկրորդը հաճախ միաձուլվում է սեպ-ուղեծրի բացվածքի հետ (for. Sphenorbitale), որը միաժամանակ սահմանափակվում է և՛ թեւ-սֆենոիդ ոսկորով, և՛ ուղեծրով։ սֆենոիդ ոսկորը ընկած է դրա դիմաց: Այս անցքով անցնում են oculomotorius III, trochlearis IV, abducens VI և trigeminal նյարդի առաջին ճյուղի նյարդերը։
Այստեղ գանգուղեղային ծածկույթը ծածկված է պարիետալ ոսկորներով (պարիետալ), որոնք հաճախ աճում են միմյանց հետ (մոնոտրեմներում, որոշ մարսոպյաններ և սմբակավոր կենդանիներ); նրանց և վերին օքսիտալ ոսկորի միջև դրված են ևս երկու ոսկորներ՝ միաձուլվելով մեկ միջպարիետալ ոսկորի մեջ (interparietale), որը որոշ կարգերում պահպանվել է որպես ինքնուրույն ոսկոր (կետասաններ, մարսուփներ, հիրաքսներ, արդվառներ, բրդոտ թեւեր և որոշ կրծողներ), մինչդեռ. Մյուսների մոտ այն աճում է միասին կամ պարիետալով (որոշ կրծողների, ծովահենների և որոճողների մոտ), կամ վերին օքսիպիտալ ոսկորի հետ (մսակեր, միասմբակավոր, պրիմատների մոտ):
Հիմնական սֆենոիդ ոսկորի առջև ընկած է առջևի սֆենոիդ ոսկորը (praesphenoideum)՝ ուղեծրային նյարդերի քիազմի իջվածքով. դրա կողերին, սովորաբար միաձուլված նրա հետ, ընկած են ուղեծրային-սֆենոիդ ոսկորները (orbitosphenoidea)՝ ուղեծրային նյարդերի անցման անցքով (for. opticum), որը կոչվում է առջևի և հիմնական սֆենոիդ ոսկորների հետ միաձուլման դեպքում, փոքր թեւեր (alae parvae): Ինչպես նշվեց վերևում, երկու pterygoids-ի միջև յուրաքանչյուր կողմում կան ճեղքաձև բացվածքներ, որոնց միջով անցնում են աչքի մկանների նյարդերը և trigeminal նյարդի առաջին ճյուղը (for. Sphenorbitale):
Ոսկրերի այս շարքի վերևում համապատասխանում են ճակատային ոսկորները (ճակատային)՝ սովորաբար լավ զարգացած զույգ ոսկորները, երբեմն միաձուլված և եղջերավոր եղջյուրների մեջ ձողեր կրող, իսկ եղջերուների մեջ՝ կանեփ եղջյուրների համար: Ծածկելով գանգուղեղը՝ ճակատային ոսկորները սահմանափակվում են ուղեծրի վերևից և ներսից՝ հաճախ ձևավորելով պրոցեսներ՝ նախաօրբիտալ (rо. Praeorbitalis և posterior-orbital (рrо. Postorbitalis): Երբեմն աչքերի շուրջը ձևավորվում է միաձուլման արդյունքում։ zygomatic ոսկոր(jugale) ճակատային ոսկրային պերիորբիտալ օղակով (որոշ միջատակեր և մսակեր, որոճողներ, միսմբակավոր կենդանիներ, կիսակապիկներ): Կապիկների մոտ ակնախորշը ժամանակավոր խոռոչից բաժանելու գործընթացն այնքան հեռու է գնում, որ ակնախորշի և ժամանակավոր խոռոչի միջև ձևավորվում է ոսկրային պատ:
Ուղեծրի առջևի պատին ընկած է արցունքաբեր ոսկոր (լաքրիմալ), որը ծակված է արցունքաբեր ջրանցքի բացվածքով (for. Lacrymale):
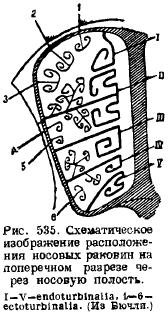
Քթի հատվածում մի քանի ոսկրացում են դրվում աճառային քթի պարկուճում։ Նախ, աճառային քթի միջնապատի երկայնքով, մեջտեղում զարգանում է ոսկրային միջնապատ (lamina perpendicularis), որը համապատասխանում է միջին հոտառական ոսկորին (mesethmoideum); երկրորդ, մեջ հետևի պատըքթի պարկուճում և ուղեղի խոռոչի առաջի պատում զարգանում են երկու կողային թիթեղներ, որոնք ծակված են բազմաթիվ [միայն պլատիպուսում այդ անցքերը փոխարինվում են մեկ (օֆակտորիումի համար)] անցքերով՝ հոտային նյարդի անցման համար՝ էթմոիդ թիթեղները (լամինա): cribrosa), որը համապատասխանում է կողային էթմոիդային ոսկորներին (ethmoidalia lateralia) ... Քթի միջնապատից ոսկրացումը տարածվում է թիկունքի կողմից կողքերի երկայնքով, անցնում քթի պարկուճների կողային պատերով և ավարտվում դրա ներքևում՝ քթի խոռոչի ներսի կողմերում ձևավորելով բազմաթիվ ելուստներ։ Սրանք հոտառական թաղանթներ են (ethmoturbinalia), որոնք երբեմն կազմում են շատ բարդ քթի լաբիրինթոս՝ կողային թիթեղների և բջջային կառուցվածքի խոռոչների հետ միասին։ Լվացարանների դիրքը երևում է կից սխեմատիկ գծագրից (նկ. 535): Այս հատվածը վերևից ծածկող քթի ոսկորից (նազալ) կախվում են քթի խոռոչի ներսում, նաև յուրաքանչյուր կողմից մեկական պատյաններ (նազոտուրբմալիա): Ներքևի վրա քթի միջնապատըչզուգակցված ոսկորը, որը նախկինում համարվում էր ձայն, համընկնում է: Այնուամենայնիվ, վերջին ուսումնասիրությունները ցույց են տվել, որ ի լրումն այս չզույգված ծածկված ոսկորին, գանգի հիմքում զարգանում են երկու զույգ ոսկորներ, որոնք կոչվում են պրևոմեր. և քանի որ սողունն ունի զույգ ոսկորի ձևով ձայն, և այս հատվածի չզույգված ոսկորը պարասֆենոիդ էր, կաթնասունների ձայնն է, որը կարելի է համարել սողունների այս վերջին ոսկորի և նախավերևանների նմանակը: համարվում են ձայնավորներ: Առջևում քթի խոռոչի պատերը ծնոտի ոսկորներն են (maxillare), որոնց վրա ձևավորվում են պատյաններ, որոնք աճում են քթի խոռոչի մեջ. սրանք ծնոտի պատյաններ են (maxilloturbinale):
Գանգի առջևի կողմն ավարտվում է միջծնոտային ոսկորների դիմաց (praemaxillare): Անթրոպոմորֆ կապիկների և մարդկանց մոտ այս ոսկորը ծնոտի ոսկորով շատ վաղ է աճում, իսկ չղջիկների մոտ այն վերածվում է փոքր ոսկորի կամ նույնիսկ ընդհանրապես բացակայում է: Իր ուղղահայաց ընթացքով միջծնոտային ոսկորը սահմանափակում է քթի բացվածքի առջևի հատվածը և տարածվում է քթի ոսկորի և ծնոտի միջև, երբեմն մինչև ճակատային ոսկորը: Ներքևից միջծնոտային ոսկորները մասնակցում են կոշտ քիմքի ձևավորմանը։ Նրանք սահմանափակում են պալատինե-պոսովյան (ստենսոն) ջրանցքի (for. Incisiva) բացվածքների կոշտ ճաշակի վրա առջևը և կողքերը, միացնելով. բերանի խոռոչՅակոբսոնի երգեհոնով։ Այս պալատինա-ռնգային բացվածքներից դուրս ընկած միջծածնոտային ոսկորների պրոցեսները վերադրվում են յուրաքանչյուր ոսկորին ինքնուրույն, դրանք, հավանաբար, սողունների զուգակցված ձայնի հոմոլոգներն են (պրևոմեր) և պահպանվում են պլատիպուսում՝ որպես անկախ ոսկորներ: Կտրիչները տեղակայված են միջծնոտային ոսկորի վրա։
Ծնոտի ոսկորները (maxillare), ինչպես արդեն նշվեց, սահմանափակում են ռնգային խոռոչի հատակը՝ հորիզոնական պրոցեսներով կոշտ քիմք կազմելով։ Քթի խոռոչի ներքին կողմում ստորին կրելի թաղանթը (maxilloturbinae) աճում է մինչև դրա կողային պատերը՝ ձևավորված ծնոտի ոսկորներից, որը դրված է որպես անկախ ոսկոր։
Կոշտ քիմքի հետընթացի շարունակությունը պալատի ոսկորների հորիզոնական պրոցեսներն են, որոնք սահմանափակում են քթի ներքին անցումը որպես կողային պատեր: Նրանք կանգնած են ուղղահայաց, և ուղղահայաց կանգնած pterygoid ոսկորները (pterygoideum) նույնպես կից են նրանց հետևում, որոնք մարդկանց մոտ միաձուլվում են սֆենոիդ ոսկորին, ձևավորելով դրա pterygoid գործընթացները (proc. Pterygoidei):
Գանգի առաջային հատվածը ենթարկվում է զգալի փոփոխությունների՝ կապված կաթեյնների, ծովահենների մոտ ջրային կենսակերպին անցնելու և փղերի բնի զարգացման հետ կապված։

Կետասանների մոտ (նկ. 536) քթանցքները շատ ետ են մղվում միջուղիղային շրջան և տանում են համապատասխանաբար ուղղահայաց ջրանցքի մեջ՝ չոաններով բացվելով բերանի խոռոչի մեջ; քթի ոսկորները ընկած են քթանցքների հետևում, վերևում դիմային ոսկորները և գրեթե միանում են միջպարիետալ ոսկորին. նաև ծնոտի ոսկորները համընկնում են ճակատային ոսկորների հետ՝ հասնելով գրեթե մինչև պարիետալ ոսկորները, որոնք միջպարիետալ ոսկորով խիստ շրջված են դեպի կողմերը։
Սիրեններն ունեն քթանցքերի նույն հետընթաց շարժումը, բայց ոչ այնքան հեռու; քթի ոսկորներն այստեղ ավելի են փոքրացել և կարող են նույնիսկ բացակայել (նկ. 537):

ժանիքների և հզոր մոլերի չափազանց զարգացած փղերի մոտ և ցողունի առկայության պատճառով, նորմալ դիրքոսկորները. քթանցքները խիստ ետ են մղվում, այս առումով քթի ջրանցքը գրեթե ուղղահայաց դիրք է գրավում. ճակատային ոսկորները շատ կարճ են, միջծնոտային ոսկորները շատ մեծ են և շատ հետ են գնում; ծնոտի ոսկորները տեղափոխվում են գանգի հիմքի տակ; ամբողջ գանգը դառնում է շատ զանգվածային՝ չափազանց զարգացած օդաճնշական ոսկորներով (նկ. 538):

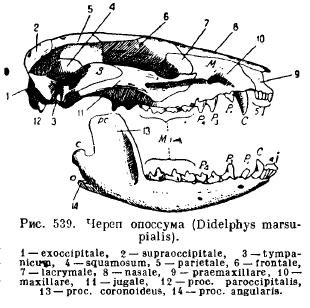
Կաթնասունների ստորին ծնոտը, ի տարբերություն մյուս բոլոր ողնաշարավորների, զարգանում է յուրաքանչյուր կողմից միայն մեկ ատամնային ոսկորից (ատամնային); երկու կեսերը կամ մնում են միմյանց հետ կապված աճառի, կարի միջոցով, կամ աճում են միասին (մեկ սմբակավոր կենդանիներ, փիղ, չղջիկներ, կապիկներ): Յուրաքանչյուր կեսը բաղկացած է հորիզոնական մարմնից (կորպուս), որի վրա նստում են ատամները, և բարձրացող մասից՝ պրոցեսներով. հետընթաց անկյունային գործընթաց (ррос . angularis); Մարսափորների մեծ մասի և որոշ կրծողների մոտ այս գործընթացը վերածվում է դեպի ներս ձգվող ափսեի (նկ. 539):
Ստորին ծնոտի նման կառուցվածքը և գանգի հետ նրա յուրահատուկ կցումը մեզ տանում են դեպի կաթնասունների այս հատկանիշների ծագման հարցը: Պատմություն անհատական զարգացումեւ պալեոնտոլոգիական տվյալները տալիս են այդ հարցի պատասխանը։ Կաթնասունների սաղմերում դրվում են պալատինե քառակուսի և մեկելյան աճառներ։ Այնուամենայնիվ, պալատին քառակուսի աճառի ոսկրացումը տեղի է ունենում ստորին ծնոտի հոդակապից դուրս և չի հանգեցնում քառակուսի ոսկորի ձևավորմանը, ինչպես սողունների մոտ, և կտա նոր լսողական ոսկոր՝ ինկուս, մինչդեռ Մեքելյան աճառը, որը ոսկրացած է իր հետևի մասում, տալիս է ոչ հոդային ոսկոր (articulare), ինչպես մյուս դիմածնոտը, իսկ երրորդ լսողական ոսկորը մալլեուսն է (malleus): Առաջին լսողական ոսկորը՝ բծերը, համապատասխանում է երկկենցաղների, սողունների և թռչունների սյունին և, ինչպես արդեն գիտենք, ձևափոխված կախոց է (hyomandibulare), այսինքն՝ հիոիդ կամարի մոտակա հատվածը։ Ճիշտ այնպես, ինչպես սյունը (columella auris), պարանոցը ծածկում է լսողական լաբիրինթոսի օվալային պատուհանը և ծակված է, ինչպես որոշ սողունների մոտ, մի անցք, որով որոշ ստորին կաթնասունների մոտ անցնում է դեմքի զարկերակը (a.facialis) (նկ. 540 և 541):

Այսպիսով, կաթնասունների մոտ քառակուսի ոսկորը վերածվել է ինկուսի, հոդային ոսկորը՝ մալլեուսի; և քանի որ փոքր ծածկված ոսկորը աճում է մինչև մուրճը, ձևավորելով դրա միջին ընթացքը, ենթադրվում է, որ այս վերջինը համապատասխանում է սողունների ծածկված հոդային ոսկորին (goniale): Այս վերջին ոսկորի կողքին ընկած՝ սողունների ծածկված անկյունային ոսկորը նույնպես անցնում է ականջի շրջան՝ դառնալով թմբկավոր ոսկոր (tympanicum): Այս փոփոխությունները տեսանելի են կից գծապատկերում (նկ. 541):
Պալեոնտոլոգիական տվյալները հաստատում են վերևում թաքնված վերափոխումների հնարավորությունը: Կենդանական ատամնավոր գազանների մեջ (Theriodontia) կան այնպիսիք, որոնց մոտ ատամնաշարը հատկապես մեծ զարգացում է ստանում՝ քառակուսի խոզանակով հասնելով ստորին ծնոտի միացման հատվածի հետնամասին։ Միևնույն ժամանակ, տափակ ոսկորը քառակուսի ոսկորով իջնում է նույն հոդակապը: Ատամնաբուժական և շերտավոր ոսկորներն այսպիսով ավելի ու ավելի են մոտենում միմյանց:
Վերջապես, որոշ կենդանիների ատամնավոր ոսկորներում ատամնաշարը և թիթեղային ոսկորները հոդային և քառակուսի ոսկորների հետ միասին մասնակցում են հոդակապմանը:
Ինչ վերաբերում է կաթնասունների հիպոգլոսային ապարատին, ապա այն բաղկացած է մարմնից, հիոիդ կամարի ձևափոխված բազալային մասից (կապուլա), առջևի եղջյուրներից (cornua anteriora), որը կազմված է հիոիդ կամարի ստորին մասից և հետևի եղջյուրներից ( cornua posteriora), վերափոխված առաջին ճյուղային կամարից։ Առջևի եղջյուրները բաղկացած են մի քանի ոսկրացումից (մինչև չորս); գանգի դեմ ուղղված հատվածը հաճախ աճում է մինչև քարքարոտ ոսկորը՝ ձևավորելով ստիլոիդ պրոցես (proc. styloideus):
Երկրորդ և երրորդ ճյուղային կամարները առաջացրել են վահանաձև գեղձի աճառի ձևավորում (cartilago thyreoidea):
Վերջույթների գոտիներ.Կաթնասունների ուսագոտին բնութագրվում է հիմնականում ագռավի ոսկորների կրճատմամբ (procoracoideum, epicoracoideum, coracoideum), ողնաշարի ոսկորների (clavicula) անհետացման հակումով և սկեպուլայի լավ զարգացած սրածայրով (spina scapulae), որը շարունակվում է դեպի մեջ: ակրոմիոն գործընթաց (ակրոմիոն):
Այնուամենայնիվ, մոնոտրեմներում ուսի գոտին մեծ նմանություն ունի սողունների հետ։ Սկապուլան հագեցած է ակրոմիալ ելուստով, որին առջևից կից է կլավիկուլը, իսկ սկեպուլայի հետևի մասում ամրացվում են ագռավի լայն և կարճ ոսկորները (coracoideum), որոնք գնում են դեպի կրծքավանդակը։ Դրանց դիմաց լայն պրոկորակոիդներ են (procoracoideum)՝ անկախ ոսկորների տեսքով։ Կլավիկուլները (clavicula) միջանկյալ միացված են էպիֆորային (episternum) (նկ. 542): Այլ կաթնասունների մոտ կրակոիդները դրվում են երկու ոսկրացման տեսքով, սակայն նրանք կորցնում են կապը կրծոսկրի հետ և աճում մինչև սկապուլա՝ ձևավորելով կորակոիդ պրոցես (proc. Coracoideus); պրոկորակոիդներն ամբողջությամբ անհետանում են (նկ. 543), գիշատիչների և սմբակավոր կենդանիների մոտ անհետանում են վզնոցները՝ արագ վազքին հարմարվելու պատճառով, իսկ կետաձկաններում և ծովախորշերում՝ առջևի վերջույթները լողակների վերածվելու պատճառով:

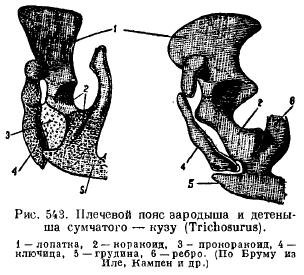
Կաթնասունների կոնքի գոտին դրված ժամանակ ունի տարրերի բնորոշ դասավորվածություն. դրա առանձնահատկությունն է, նախ, լրացուցիչ ոսկորի առկայությունը ացետաբուլարում (acetabulare), և երկրորդը, այս բոլոր ոսկորների միաձուլումը յուրաքանչյուր կողմից մեկ անանուն ոսկորի մեջ (innominatum) և միաձուլվելը ներքևի և իշիալ ոսկորների միջև, այնպես որ. փակ հոդ է ձևավորվում այս ոսկորների միջև անցքի բոլոր կողմերից (համար. Obturatum) (նկ. 544): Մոնոտրեմների և մարսուալների մոտ ներքևից հատուկ ոսկոր է ամրացվում ներքևից՝ պարկը պահելու համար (marsupiale):
Վերջույթների կմախքկաթնասունների սկզբնական ձևերում դա պայմանավորված է արագ շարժումներին նրանց հարմարվողականությամբ՝ վազում չորս վերջույթների օգնությամբ, որոնք գտնվում են մարմնի տակ, որպեսզի վերջիններս կախված չլինեն վերջույթների միջև, ինչպես սողունների մեծ մասում, այլ հանգստանա։ ուղղակիորեն նրանց վրա: Վազելու ունակությունը պայմանավորված է կաթնասունների մոտ վերջույթների տարբեր դիրքով։ Նրանց հետ է, որ վերջույթները կողմնորոշվում են այնպես, որ առջևի վերջույթների արմունկների հոդերը ուղղվեն դեպի հետ, իսկ հետևի ծնկների հոդերը՝ առաջ։ Շարժվելիս առջևի վերջույթները նետվում են առաջ և քաշում մարմինը՝ դաստակը թեքելով և արմունկի միացումներ; հետևի վերջույթները մղում են մարմինը առաջ՝ ուղղելով ոտքը ծնկահոդի մոտ:
Ըստ ոսկորների քանակի, որոնք կազմում են վերջույթները և ըստ իրենց գտնվելու վայրի, կաթնասունների նախնիների ձևերի վերջույթների կմախքը ունի պարզունակ հինգ մատով վերջույթի շատ բնորոշ կառուցվածք և ենթարկվել է. մեծ փոփոխություններդասարանի ներսում միայն տարբեր տեսակի շարժումներին հարմարվելու գործընթացում տարբեր պայմաններչորեքշաբթի.
Առջևի վերջույթը բնութագրվում է դիստալ շրջանում առկայությամբ humerusանցքեր (for.entepicondylare), ulna-ի պրոքսիմալ ելուստի առկայություն (olecranon), ցողունային ոսկորի առկայություն (pisiforme) և չորրորդ և հինգերորդ հեռավոր կարպալ ոսկորների միաձուլում (carpalia dis talia 4 + 6 = hamatum, կամ uncinatum) և կենտրոնական պրոքսիմալ միջանկյալ հետ (centra dist. + intermedium = lunatum, կամ semilunare): Շնորհիվ այն բանի, որ մարդու անատոմիայում ընդունված կաթնասունների դաստակի ոսկորների նոմենկլատուրան հաճախ օգտագործվում է, մենք այս անունների աղյուսակը տալիս ենք դիագրամում, որը ցույց է տալիս ոսկորների գտնվելու վայրը:
Կաթնասունների հետևի վերջույթը հիմնականում բնութագրվում է նրանով, որ ազդրը ծնկահոդով պտտվում է ազդրի առջևի մասում և ունի լավ զարգացած պրոցեսներ պրոքսիմալ մասում՝ տրոհանտերներ՝ թվով 2-3 (տրոհանտեր մաժոր, մինոր և տերցիուս): ); սրունքում սրունքն ավելի զարգացած է, կա ոսկրային պաթելլա (պաթելլա), որը պաշտպանում է ծնկահոդը։ Տարսուսում կան միայն երկու մոտակա ոսկորներ՝ լավ զարգացած՝ ներքինը՝ թալուսը (talus = astragalus), որը ձևավորվել է միջանկյալ և կենտրոնական պրոքսիմալի միաձուլումից (intermedium + centrale proximale) և արտաքին calcaneus (calcaneus), համապատասխանում է թարսուսի ֆիբուլյար ոսկորին (ֆիբուլյար): Միջին շարքի ոսկորներից կենտրոնական դիստալը (centrale distale) վերածվել է սկաֆոիդի (naviculare = scaphoideum): Ոտնաթաթի ոսկորը կա՛մ կրճատվել է, կա՛մ սոսնձվել է այս վերջինիս վրա:
Տարսոնի և մետատարսուսի ոսկորների դասավորությունը և անունների հոմանիշները հետևյալն են.
Տարբեր միջավայրերում տեղաշարժին հարմարվելու հետ կապված՝ կաթնասունների վերջույթները ենթարկվել են շատ զգալի փոփոխությունների, և այդ փոփոխությունները հատկապես լավ կարելի է նկատել կմախքի վրա։ Ցավոք, մենք չունենք պալեոնտոլոգիական տվյալներ կաթնասունների նախնիների վերջույթների կառուցվածքի վերաբերյալ։ Կասկած չկա, որ առաջին կաթնասուններն ունեին վերը նկարագրված տիպի քիչ մասնագիտացված վերջույթներ: Սրանք ժամանակակից կաթնասունների ամենապրիմիտիվ մարսուալների վերջույթներն են, օրինակ՝ օպոսումները՝ մարսուալ առնետները (Դիդելֆիս) և միջատակերները, որոնք վարում են ծառի կենսակերպ: Այնուամենայնիվ, սխալ կլինի դրանից եզրակացնել, որ կաթնասունների նախնիները, անշուշտ, անտառների և առավել եւս ծառերի վրա բարձրացող ձևերի բնակիչներն են եղել։ Բացի այդ, պոզումի վերջույթը, թեև այն ծառայում է ծառերի ճյուղերի երկայնքով շարժվելու համար, ամենևին էլ տիպիկ մասնագիտացված ծառի վերջույթ չէ, բայց պահպանում է իր ունիվերսալ հնգմատանի տեսակը։ Նման վերջույթները լավագույնս կարող էին գոյատևել այն կենդանիների մոտ, որոնք ոչ միայն վազում էին գետնին, այլև ծառեր էին մագլցում, ինչպես նաև այն կենդանիները, որոնք լեռներ էին մագլցում: Կաթնասունների վերջույթների մասնագիտացումը ընթացավ միաժամանակ կարգերի և այլ փոքր համակարգային միավորների տարբերակման հետ, և տեղի ունեցավ սերտաճում։
Վերջույթները ենթարկվում են հատկապես խորը փոփոխությունների, երբ հարմարվում են թռիչքին, ջրում լողալուն, գետնի տակ շարժվելիս. հողի դանդաղ շարժումները և ծառաբնակ ապրելակերպը կապված չեն խորը մասնագիտացման հետ:
Չղջիկների մոտ (Chiroptera) առջևի վերջույթը վերածվում է թևի, իսկ օդային փայլաթիթեղը ձգվում է չորս (2-բ) խիստ երկարացած մատների միջև՝ առանց ճանկերի. առաջին մատը կարճ է և վերջում ունի ուժեղ զարգացած ճանկ; այս մատը ոչ մի դեր չի խաղում թռչելու մեջ, այն օգտագործվում է մագլցելու համար; դաստակում, սկաֆոիդը, լուսնային և կենտրոնական ոսկորները միասին աճում են մեկ; նախաբազուկը երկարաձգված է և կազմված է լավ զարգացած շառավղից, իսկ ուլնան փոքրացած է և ավելանում է շառավղով: Օդային փայլաթիթեղը կլանում է նաև հետևի ոտքերը: Վերջիններիս միջև այն տարբեր կերպ է զարգացած չղջիկների մոտ (նկ. 545):
Ջրի մեջ լողացող կաթնասունների մոտ վերջույթները վերածվում են լողակների այնպես, ինչպես մենք արդեն տեսել ենք սողունների մոտ։ Ընդ որում, հատկապես փոխվել են կետերի վերջույթներն ու ազդանշանները։ Ի տարբերություն թևի, առաջնային վերջույթն իր մոտիկ հատվածում զգալիորեն կրճատվել է։ Կետերի մոտ ուսի և նախաբազկի ոսկորները վերածվել են կարճ, լայն թիթեղների, որոնք տեղ են ապահովում հզոր մկանների ամրացման համար: Վերջույթի բոլոր մասերը անշարժ կապված են միմյանց հետ՝ կազմելով ամուր թիակ։ Այս դեպքում ֆալանգների թիվը կարող է մեծապես աճել: Նույն փոփոխությունները տեսանք սողունների մոտ։ Ֆալանգների թվի աճը կախված է նրանից, որ ֆալանգների դիաֆիզը չի աճում սոճու գեղձերի հետ միասին և ոսկրանում է առանձին կենտրոններից։ Նման առանձին ոսկրացումը պայմանավորված է այս կենդանիների ոսկրացման հետաձգված գործընթացով։ Ոսկրացման գործընթացի դանդաղությունը հանգեցնում է նրան, որ իրենց հանգույցում գտնվող ոսկորների մեծ մասը մնում է աճառ:
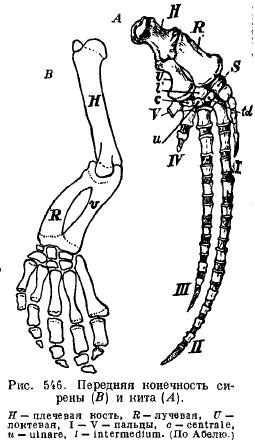
Սիրենների լողակները (Sirenia) արտաքին նմանություն ունեն կաթեյնիկների լողակների հետ. ուսն ու նախաբազուկը միմյանց հետ հոդակապված են, որպեսզի կարողանան թեքվել. կան դաստակի գրեթե բոլոր ոսկորները և բոլոր հինգ մատները՝ մի փոքր փոփոխված ֆալանգների քանակով (նկ. 546):
Հետևի վերջույթները ներս ժամանակակից կետերիսկ ազդանշանները կրճատվում են այնպես, որ դրանցից մնում են միայն կոնքի մնացորդները (նկ. 547 և 548):
Վերջույթների օջախում շարժմանը հարմարվելու մեկ այլ տեսակ մենք տեսնում ենք պտուտակավորների մոտ (Pinnipedia): Այստեղ պոչը գրեթե անհետանում է, իսկ հետևի ոտքերը հետ են շարժվում՝ դեր խաղալով շարժիչ օրգան... Այս կարգի երեք ընտանիքները, ըստ ջրի շարժման հարմարվողականության զարգացման աստիճանի, ներկայացնում են երեք փուլ, ինչպես դա եղել է (ոչ գենետիկորեն): Ծովերի և ականջավոր փոկերի մոտ (Otariidae) հետևի վերջույթները դեռ կարող են թեքվել դեպի առաջ մարմնի տակ, իսկ ծովացուլերի մոտ վերջույթների կառուցվածքում ավելի պարզունակություն կա, դրանք ավելի կարճ են և ավելի շարժուն։ Փոկերի մոտ (Phocidae) հետևի ոտքերը այլևս չեն կարող առաջ թեքվել մարմնի տակ, սակայն մագիլները դեռևս պահպանվում են մատների վրա (ֆլիպեր):
Փորող կենդանիների մոտ վերջույթները նույնպես ենթարկվում են շատ ուժեղ մոդիֆիկացիաների. հատկապես ուժեղ են փոխվել մարսապի խալերի (Notoryctes typhlops) վերջույթները։ Նա ուսի շեղբին ունի երկու սրածայր, ինչպես շատ փորող կենդանիներ, ինչը պայմանավորված է ուսի եռգլուխ մկանների ուժեղ զարգացմամբ. ունի վզկապ, ինչպես բոլոր փորված կաթնասունները: Բազուկն ունի շատ ամուր եզր, որը ճյուղավորվում է պատառաքաղանման ձևով և ծառայում է ամուր (փորելու համար անհրաժեշտ) դելտոիդ և կրծքային մկանները; ուսի ներքին կոնդիլը բարձր զարգացած է շնորհիվ ուժեղ զարգացում ulna-ի ճկուն մկանները (proc. olecranon), որը ծառայում է որպես triceps մկանների կցման կետ; դաստակը յուրօրինակ փոխված է, դառնում է շատ ամուր; 1-ին և 2-րդ մատները հետ են մղվում դեպի ափի կողմը, և միայն 3-րդ և 4-րդը, որոնք հագեցած են հսկայական ճանկերով, հատկապես 3-րդը, ծառայում են փորելու համար. մատների մոտ ֆալանգների թիվը շատ փոքր է. 2 և 1. 5-րդ մետակարպալ լայն հարթ ճանկով, որը մեծացնում է փորվածքի մակերեսը; ափի հարթությունում կա քսամոիդ ոսկոր, որին կցվում է մկանը՝ թեքելով մատները, որը մեծ դեր է խաղում փորելու մեջ. այն մեծացնում է դաստակի ուժը; ինչպես մյուս փորվածքային ձևերը, սիմֆիզը pubic ոսկորներշատ կարճ; Իշիումը աճում է երկու վերջին սրբային ողերի լայնակի պրոցեսների հետ միասին. ազդրի, ծնկի և սրունքի վրա բարձր զարգացած են ծայրերն ու ելքերը՝ մկանների կցման տեղերը. սրունքը և սրունքը (սրունքը և թմբիկը) ամուր միաձուլված են, ինչպես ոսկե խլուրդ (Chrysochloris); ներբանը ծալված է դեպի դուրս, ինչպես Chrysochloris-ում; 5-րդ մատը թեքված է ոտքի մակերևույթի վրա, ոտքի 5-րդ եղունգը լայն է և թիակաձև, 4-րդը երկար է և մանգաղաձև, 3-րդ և 2-րդը դառնում են ավելի կարճ և լայն. կրծքավանդակի վրա ուժեղ զարգացած գագաթ է, ինչպես ոսկե խալը, խալ առնետը (Spalax) և խալը (Talpa) (նկ. 549):
Ոսկորների վրա մկանների ամրացման համար սրածայրերի ուժեղ զարգացում, կարպուսի կարճացում, կարպալիայի ամուր կապ, որոշ ֆալանգների կրճատում և փորելու մեջ դեր ունեցող մյուսների ուժեղացում. այս բոլոր նշանները բնորոշ են. փորող կաթնասուններ ընդհանրապես. Բայց կախված նրանից, թե կենդանին ինչ հող է փորում, տարբերություններ կլինեն։ Փափուկ հողը փորելիս (օրինակ՝ խլուրդում՝ Թալպա) (նկ. 550) ձեռքը մեծապես լայնանում է, գործում է թիակի պես և հակված է զարգացնել ոտքի 6-րդ մատը։ Խլուրդի առջևի ծայրը համեմատելի է փափուկ հողում փորելու համար օգտագործվող թիակի հետ։ Ընդհակառակը, այն ձևերում, որոնք փորում են կոշտ հողում, ձեռքը նեղանում է, փորելուն մասնակցում են միայն մի քանի մատներ, հիմնականում 3-րդ և 4-րդը, զարգանալով ավելի ուժեղ, քան մյուսները, ինչպես Notoryctes-ում և Chrysochloris-ում։ Այս խոզանակը համեմատելի է կոշտ հողի փորման բարձիկի հետ:
![]()
Նույնը մենք տեսնում ենք կաթնասունների մոտ, որոնք պատռում են տերմիտների պինդ կառուցվածքները, օրինակ՝ մրջնակեր (Myrmecophagus), տամանդուայում (Tamandua): Այս կենդանիները քայլում են՝ հենվելով ոտքերի արտաքին եզրին, ճանկերը ծալած։
Կաթնասունների հարմարեցումը գետնի վրա արագ շարժմանը շատ էական փոփոխություններ է առաջացրել վերջույթներում։ Գետնի վրա շարժումը, որը բնորոշ էր պարզունակ կաթնասուններին, իրականացվում էր ամբողջ ձեռքի և ոտքի վրա հենվելով; բնօրինակ ձևերԿաթնասունները հիմնականում ցողունային կամ ցողունային էին, այսինքն՝ քայլելիս հենվում էին ամբողջ ոտքերի վրա, ինչպես, օրինակ, արջերը։ Բայց նման կառուցվածքը թույլ է տալիս միայն համեմատաբար դանդաղ քայլել՝ համեմատած սրընթաց ոտքով անտիլոպների կամ ձիերի, կամ թռչկոտող նապաստակների, կենգուրուների, ջերբոների հետ: Ֆիլոգենետիկ զարգացման ընթացքում մենք տեսնում ենք դաստակի և տարսոնի գետնից աստիճանական բարձրացում (կարպուս և թարսուս), որին հաջորդում են մետակարպուսը և մետատարզուսը, բայց ֆալանգները դեռ ամբողջությամբ դիպչում են գետնին: Ի վերջո ուղղվեցին նաեւ ֆալանգները, առջեւի եւ հետեւի վերջույթները սկսեցին հենվել մատների ծայրերին։ Այժմ մենք կարող ենք քայլ առ քայլ հետևել այս էվոլյուցիայի փուլերին ոչ միայն սմբակավոր կենդանիների, այլև մսակեր կաթնասունների մոտ, որոնք արագ վազելու ունակություն են ձեռք բերել: Երբ վերջույթները ուղղվում են, ծայրահեղ մատները դադարում են դիպչել գետնին, դադարում են գործել և ենթարկվում են ատրոֆի: Արդյունքը գետնի հետ շփման ավելի փոքր մակերես է: Վերջույթների հեռավոր մասի փոփոխությունները հանգեցրին պրոքսիմալ փոփոխություններին. կարպալիան և տարսալիան ավելի ու ավելի սերտորեն տեղակայված են միմյանց կողքին, ուլնան և ֆիբուլան կրճատվում են, վերջույթները սկսում են շարժվել միայն սագիտալ հարթությունում, միջնադարյան մետապոդիան միաձուլվում է. մեկ ոսկոր.
Զարգացման երկու ուղիները հանգեցնում են ժամանակակից սմբակավոր կենդանիների վերջույթների ձևավորմանը. Մի շարքով վերջույթի առանցքն անցնում է երրորդ մատի միջով և վերջինս զարգանում է ամենաուժեղ, իսկ մնացածները դառնում են տարրական։ Սրանք էկվիդներ են (Perissodactyla), մեսաքսիալ (Mesaxonia): Մյուսներում առանցքը անցնում է երրորդ և չորրորդ մատների միջև՝ զարգանալով հավասարաչափ՝ արտիոդակտիլներ (Artiodactyla), պարաքսիալ (Paraxonia): Կողային մատները կրճատվում են: Էկվիդների զարգացումը առավել հստակ երևում է ձիու վերջույթների ֆիլոգենետիկ զարգացման մեջ (նկ. 551): Սմբակավոր վիճակի զարգացումը, ի թիվս այլ բաների, պայմանավորված էր նրանով, որ դրան նախորդել էր մատի առաջին մատի փոքրացումը, այնպես որ այն գրեթե չի մասնակցել ոտքով քայլելուց անցմանը, մինչդեռ նախնիների մոտ. միասմբակով, հինգերորդ մատի զարգացումը սկզբում նորմալ էր:
Հետաքրքիրն այն է, որ և՛ սմբակավորությունը, և՛ կենտ սմբակները առաջացել են էվոլյուցիոն գործընթացում բազմիցս և որոշ դեպքերում հանգեցրել են շատ մասնագիտացված տեսակի, օրինակ՝ հարավամերիկյան Proterotlieriidae-ում (անհետացած):
Մնացած մատները պատված են սմբակներով, որոնք կարող են ավելի տարբերվել՝ կախված ապրելակերպից և բնակավայրից: Օրինակ՝ ժայռերից բռնած այծերն ունեն կտրուկ և նեղ սմբակներ՝ սուր ծայրով. ժամը հյուսիսային եղջերուդրանք շատ երկար են, լայնորեն երկատված և ծառայում են որպես մի տեսակ դահուկներ, որոնք թույլ չեն տալիս կենդանուն ընկնել ձյան և ճահիճների մեջ:
Մատների հետ մատների վրա քայլելու զարգացման հետ միաժամանակ տեղի է ունենում դաստակի (կարպուս) և թարսուսի (տարսուս) փոփոխություն։ Այն տարբեր կերպ է ընթանում արտիոդակտիլներում և էկվիդներում՝ ձգողականության տարբեր բաշխման պատճառով: Վերջապես, հետզհետե քայլվածքին անցնելու հետ կապված, ուլնայի արժեքը՝ որպես ուսի հենարան, աստիճանաբար նվազում է՝ այն նվազում է կամ միաձուլվում ճառագայթային (շառավղով) հետ։ Նմանապես, ֆիբուլան կրճատվում է և վերջապես անհետանում է հետևի վերջույթներում:
Նկարագրված փոփոխությունների իմաստն ու բնույթը պարզ երևում են ուղտի և փղի վերջույթների համեմատությունից (նկ. 552): Փղի մոտ ձեռքը (ոտքը) մնում է կարճ, իսկ ուսն ու նախաբազուկը (ազդր և ստորին ոտք) երկարացված են։ Ընդհակառակը, ուղտի մեջ ակրետային մետապոդիաները երկարացվում են. ձեռքի վերին և նախաբազուկը (ինչպես նաև ազդրը և ստորին ոտքը) մնում են կարճ: Մի շարք ոսկորների կորստի պատճառով վերջույթի կմախքը պարզեցվում է, վերջինս դառնում է ավելի թեթեւ։ Մատների կրճատումը հանգեցնում է ավելի քիչ շփման, նվազում է վնասվածքի, կապանների պատռման վտանգը։ Metapodia-ի միաձուլումը մեծ ուժ է ստեղծում: Փղի վերջույթը շատ ավելի ծանր է։ Ձեռքի ոսկորների առատությունը պահանջում է բոլոր մատների միացում՝ կայունություն ձեռք բերելու համար։ ընդհանուր մաշկ... Փղի վերջույթը մեծ ամրության սյուն է, բայց զուրկ է այն թեթևությունից, որ ունի ուղտի վերջույթը։
Գետնի վրա արագ շարժվելու մեկ այլ միջոց է ցատկել: Այս ադապտացիան բազմիցս հայտնվել է կաթնասունների պատմության մեջ և, առավել ևս, ֆիլոգենետիկ կոճղերի լայն տեսականիում՝ մարսապների շրջանում՝ կենգուրուներում (Macropus), միջատակերների մոտ՝ թռչկոտողների մոտ (Macroscelidae), և վերջապես, տարբեր աստիճաններկրծողների զարգացում. սա. Նապաստակներ (Leporidae) Duplicidentata-ից; խոզապուխտների շարքում (Hystricomorplia) զարգացել է մի խումբ striders (Pedetinae); մկնանման (Մյոմորֆա) տեսակներից մենք ունենք ավստրալական ջերբոա (Conilurus), Հին աշխարհի ջերբոա (Jaculus, Alactaga), ամերիկյան ցատկողներ (Zapus) և ցանցաթաղանթներ (Dipodomys):
Ցատկելու ունակության զարգացումը կապված է հետևի վերջույթի, այն է՝ մետատարսուսի (մետատարսուս) ոսկորների երկարացման հետ։ Բացի այդ, դա հանգեցնում է ավելորդ էքստրեմալ մատների կորստի, և թարսուսի ոսկորները շատ ամուր կապված են միմյանց հետ: Օրինակ՝ հողեղեն նապաստակի (Ալակտագա) և իսկական ջերբոայի (Դիպուս) ոտքերը։
Ալակտագայի առջևի ոտքերը փոքր են և կարճ: Հողային նապաստակն օգտագործում է դրանք փորելիս, բայց ցատկելիս չի դիպչում գետնին։ Թռիչքն իրականացվում է միայն հետևի ոտքերի օգնությամբ և նույնիսկ առանց պոչի օգնությամբ։ Վերջինս գետնին դիպչում է միայն հանգիստ վիճակում, երբ կենդանին հենվում է պոչին։ Ոտքի կմախքն ունի այն հատկանիշը, որ 2-րդ, 3-րդ և 4-րդ մատների մետատարսային ոսկորները միաձուլվում են մեկ ոսկորի մեջ։ Ներքևում գտնվող մետատարսային ոսկորների ծայրերը միաձուլված չեն, և միջնամատը (3-րդ մատը) դուրս է ցցվում ավելի առաջ: Նրանց հետ կապված ֆալանգներից միջինները կրկին ամենաերկարն են։ 1-ին և 5-րդ մատները տարրական են և չեն դիպչում գետնին: Այս երկու մատներն էլ ունեն հավասար երկարությունպայմանավորված է նրանով, որ metatarsal ոսկոր բութ մատը 5-ից ավելի երկար։ Առաջինն ունի 2, երկրորդը՝ 3 ֆալանգ։
Դիպուսում մասնագիտացումն էլ ավելի հեռուն գնաց. 1-ին մատն անհետացավ, 5-ից միայն ռուդիմենտ է մնացել։ Միջնամատերկու անգամ ավելի թույլ, քան երկու ծայրահեղները, և միայն մի փոքր ավելի երկար, քան նրանց: Ոտքի մատները զուգահեռ են: Միջնամատը կրճատվում է, քանի որ նրա գործառույթը ստանձնում են 2-րդ և 4-րդը, որոնց ծայրերում շեշտը դրվում է նստելիս և ցատկելիս։ Հետագա էվոլյուցիայի ժամանակ 3-րդ մատը պետք է անհետանա. Դիպուսում ունենք ադապտացիայի անավարտ տեսակ։