З чого складається грудний відділ хребта ссавців. Зовнішня будова ссавців, скелет та м'язи на прикладі домашньої собаки
Хребет складається з шийного, грудного, поперекового, крижового та хвостового відділів. Його характерна особливість- платильна (з плоскими поверхнями) форма хребців, між якими розташовані хрящові міжхребцеві диски. Добре виражені верхні дуги. У шийному відділі є сім хребців, від довжини яких залежить довжина шиї; тільки у ламантина і лінивця - Choloepus hof-fmani їх 6, а у лінивця- Bradypus 8-10. Шийні хребці дуже довгі у жирафа і дуже короткі у китоподібних, які не мають шийного перехоплення. До хребців грудного відділу причленовуються ребра, що утворюють грудну клітку. Грудина, що замикає її, плоска і тільки у кажаніві риючих видів з потужними передніми кінцівками (наприклад, кротів) має невеликий гребінь (кіль), що є місцем прикріплення грудних м'язів. У грудному відділі 9-24 (частіше 12-15) хребців, останні 2-5 грудних хребців несуть -хибні ребра-, що не доходять до грудини., У поперековому відділі від 2 до 9 хребців; зі своїми великими поперечними відростками зливаються рудиментарні ребра. Крижовий відділ утворений 4-10 зрослими хребцями, з яких лише два перші істинно крижові, а решта - хвостові. Число вільних хвостових хребців коливається від 3 (у гібона) до 49 у довгохвостого ящера.
Різна міра рухливості окремих хребців. У дрібних бігаючих і лазаючих звірів вона велика, по всій довжині хребта, тому їхнє тіло може згинатися в різних напрямкахі навіть згортатися в клубок. Менш рухливі хребці грудного і поперекового відділів у великих тварин, що швидко рухаються. У ссавців, що пересуваються на задніх нігах (кенгуру, тушканчики, стрибунчики), найбільші хребці перебувають у основі хвоста і крижів, а далі вперед їх величина послідовно зменшується. У копитних, навпаки, хребці і особливо їх остисті відростки більші в передній частині грудного відділу, де до них прикріплюється потужна мускулатура шиї та частково передніх кінцівок.
Зовнішня будова собаки
Собака був першою домашньою твариною. Людина приручила її ще в давнину. Собака допомагав первісній людині під час полювання, охороняв його житло. Наразі відомі собаки службові, мисливські та декоративні, а також дворняги (безпородні). Собаки допомагають нести службу з охорони кордонів нашої Батьківщини, а під час Великої Вітчизняної війнивідшукували поранених, допомагали у розвідці та зв'язку. На собаках вчені проводять дослідження життєдіяльності організму. Собака раніше за людинупобувала у космосі.
Тулуб собаки, наприклад вівчарки або лайки, стрункий і м'язистий. Її кінцівки розташовуються не з боків тіла, як у плазунів, а під тулубом, тому тіло тварини не стосується землі. Ходить собака, спираючись на пальці з міцними кігтями. Гнучка шия сприяє великій рухливості голови. Рот тварини обмежений рухомими губами - верхньою та нижньою. Над верхньою губоюзнаходиться ніс із парою зовнішніх носових отворів - ніздрів. Очі мають добре розвинені повіки. Миготлива перетинка (третя повіка) у собаки, як і у всіх ссавців, недорозвинена. З усіх тварин тільки у ссавців є зовнішнє вухо - вушна раковина. Зовнішнє вухо у собаки велике та рухоме.
Покрови ссавців
Шкіра ссавців міцна і еластична, у більшості звірів у ній містяться основи волосся, які утворюють характерний для цього класу хребетних. волосяний покрив . Розрізняють товсті та довге волосся — істьі коротші, м'які - підпуш, або підшерстя. Груба та міцна остюжка захищає підшерстя та шкіру від пошкоджень. Підшерстя, в якому затримується багато повітря, добре зберігає тепло тіла. Крім ості та підпуші у звірів розвиваються велике волосся- Органи дотику.
Волосся ссавців, подібно до пір'я птахів і луски плазунів, складаються з рогової речовини. Два рази на рік собака, як і багато інших ссавців, линяє - частина її волосся випадає і замінюється новим. Основа волосся розташована всередині особливої волосяної сумки, куди відкриваються протоки, що знаходяться поруч. сальних залоз. Їх виділення змащують шкіру і волосся, через що вони набувають еластичність і не змочуються водою.
Малюнок: Будова шкіри та волосся ссавця
У шкірі більшості звірів розташовані і потові залози . Піт, випаровуючись із поверхні тіла, охолоджує його. Разом з згодом з тіла виводиться також надлишок солі та сечовини; таким чином, потові залози відіграють роль додаткових органіввиділення.
У собаки потових залозмало та охолодження організму досягається при почастішанні дихання.
На кінцях пальців ссавців знаходяться рогові пазурі, нігті або копита. Іноді рогові утворення розвинені також на голові (роги у носорогів, антилоп, рогатої худоби та ін) або на хвості (наприклад, рогові лусочки у щурів).
Скелет ссавців
Скелет ссавців складається з тих самих відділів, що і в інших хребетних. Череп звірів відрізняється більшою черепною коробкою, що пов'язано з великими розмірамиголовного мозку. Для ссавців дуже характерною є наявність 7 шийних хребців. І у довгоший жирафа, і у китів число шийних хребців однаково. Грудні хребці (зазвичай їх 12-15) разом із ребрами та грудиною утворюють міцну грудну клітину. Масивні хребці поперекового відділурухомо зчленовані між собою. У цьому відділі тулуб може згинатися та розгинатися. Число поперекових хребців різне у різних видів(2-9), у собаки їх 6. Крижовий відділ хребта (3-4 хребця) зростається з кістками тазу. Число хребців хвостового відділу (від трьох до кількох десятків) залежить від довжини хвоста.

Малюнок: Скелет ссавців на прикладі собаки

Малюнок: Череп собаки
Пояс передніх кінцівокссавців складається з двох лопаток з воронячими кістками, що приросли до них, і двох ключиць. У собаки ключиці не розвинені. Пояс задніх кінцівок- таз - утворений трьома парами тазових кісток. Скелети кінцівок у різних видів ссавців і плазунів подібні, проте деталі їх будови у різних видів різні і залежать від умов проживання тварини.
М'язи ссавців
У більшості ссавців, у тому числі й собаки, особливо розвинені м'язи спини, кінцівок та їх поясів. Собака може бігти великими стрибками, згинаючи та розпрямляючи тулуб, по черзі відштовхуючись від землі то передніми, то задніми ногами. Сильні м'язи надають руху нижню щелепу. Стисаючи щелепи, озброєні зубами, собака міцно утримує видобуток.

Малюнок: М'язи собаки, що рухають нижню щелепу
Плечовий пояс ссавців спрощений, складається з великої лопатки, що має коракоїдний виріст, та ключиці(Рис. 80, 81). На ембріональній стадії розвитку та у дорослих однопрохідних звірів коракоїдє самостійною кісткою.
Лопаткарозташована поверх грудної клітки, має трикутну форму і високий виріст (гребінь) акроміальнимвідростком для прикріплення м'язів. У місці розташування коракоїдного виросту та акроміального відростка лопатки знаходиться суглобова ямкадля причленування передньої кінцівки.
Ключицяпаличкоподібної форми служить для з'єднання плеча та грудини. Найбільш розвинена ключиця у тварин, що виконують передні кінцівки. складні рухи(Кажани, кроти, примати, гризуни та ін.). Багато видів, котрим характерні одноманітні руху (копитні, собачі), ключиця відсутня. В цілому пояс передніх кінцівок пов'язаний з осьовим скелетом за допомогою м'язів і зв'язок.
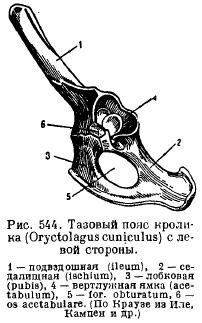
Тазовий пояс (Рис. 82 А) складається з парних безіменних кісток, утворених зчленуванням клубових, сідничних та лобковихкісток. Таз закритого типуі через здухвинні кістки міцно з'єднаний з крижовим відділомхребта. При зрощенні елементів тазу виникає лонне зрощеннята формується вертлюжна западинадля причленування задньої кінцівки. У нижній частині безіменної кістки є замикальний отвір.
Скелет парних кінцівок
Кінцівки ссавців, як і в інших наземних хребетних, п'ятипалого типу і є тричленним важелем, всі частини якого рухомо з'єднані між собою.
Передня кінцівка складається з плеча, передпліччя та кисті(Рис. 81). Плечова кісткадобре розвинена, має округлу голівку для причленування до пояса кінцівки. Передпліччяскладають дві кістки – променева та ліктьова, що має ліктьовий відростоку верхній частині. У місці з'єднання плеча та передпліччя знаходиться ліктьовий суглоб.
Пензлик включає три відділи – зап'ястя, п'ясть та фаланги пальців. Зап'ястіскладається з 8-10 елементів, розташованих у три ряди . П'ястьвключає п'ять кісток (за кількістю пальців). Кожен палець має три фаланги, крім першого пальця, що з двох фаланг. На відміну від інтеркарпального зчленування рептилій та птахів, рухливий суглобу ссавців розташований між передпліччям та пензлем(як у земноводних).
Задня кінцівка ссавців також має три відділи – стегно, гомілка та стопа(Рис. 82 Б). Стегнова кісткамасивна і вставлена округлою головкою у вертлюжну западину тазового пояса. Гомілка включає дві кістки – велику гомілкову і малу гомілкову.Більша великогомілкова кістка становить внутрішній елемент гомілки, а малогомілкова кістка – її зовнішню частину.
Між стегном і гомілкою є суглоб, прикритий колінним філіжанкою, яка еволюційно сформувалася їх окостенілим м'язовим сухожиллям. Поверхні кісток, що зчленовуються, покриті хрящовими утвореннями. Гомілковостопний суглобссавців розташований між нижніми кінцями гомілкових кісток гомілки та проксимальним відділом стопи.

Мал. 82. Тазовий пояс (А) та задня кінцівка (Б) лисиці:
1 – клубова кістка, 2 – сіднична кістка, 3 – лобкова кістка, 4 – вертлюжна западина, 5 – стегнова кістка, 6 – колінна чашка, 7 – велика гомілкова кістка, 8 – мала гомілкова кістка, 9 – передплюсна, 10 – плюсна, 11 – фаланги пальців, 12 – таранна кістка, 13 – п'яткова кістка.
Передплюсневі елементистопи розташовані в три ряди і утворюють виріст – п'яткову кістку, характерну для ссавців. Плюснаскладається з п'яти кісток, яких причленяются фаланги пальців. Як і в передніх кінцівках, їх по три у кожному пальці, крім першого (внутрішнього), який має лише дві фаланги.
Для ссавців характерна особлива постановка кінцівок щодо тіла: вони розташовані під тулубом, що сприяє розвитку більшої маневреності рухів. При цьому ліктьовий суглоб спрямований дозаду, а колінний суглоб, Навпаки, орієнтований допереду.
У зв'язку з видовим різноманіттям ссавців, їх способом життя і способами пересування значно варіюють відносна довжина кінцівок, співвідношення складових елементів, конфігурація кісток, число пальців тощо. службовців для прикріплення літальної перетинки.
Навпаки, передня кінцівка крота вкорочена, розширена долонна частина кисті та добре розвинений шостий палець. У китів різко подовжена кисть за рахунок збільшення числа фаланг пальців, внаслідок чого кінцівка набула велика подібністьз плавником кістеперих риб. Значно редуковані та видозмінені пальці тварин, що володіють великою біомасою та швидким бігом (непарнокопитні, парнокопитні).
Однією з особливостей скелета ссавців є великий розвитокхряща у молодих тварин та тривале збереження його у постембріональний період розвитку. Це пояснюється особливостями зростання хряща і кістки у зв'язку зі зростанням самої тварини. Саме хрящ росте всією своєю масою (інтерстиціально), зростання відбувається швидко і пропорції частин при цьому можуть зберігатися, тоді як кістка росте повільніше і тільки з поверхні, так що частини скелета, що з'явилися, залишаються завжди на попередній відстані і збереження пропорції частин при зростанні кістки виявляється неможливим. Ось чому молоді тварини завжди містять багато хряща і особливо у таких місцях, де сильно здійснюється зростання. Так, довгі кісткиКінцівки завжди складаються з середньої частини - діафізи, яка окостенює насамперед, і кінців, які довго зберігаються хрящовими - епіфіз. Окостеніння діафізи починається насамперед з периферії - перихондрально, тобто за рахунок клітин, що походять із сполучної тканини, що одягає хрящ, - з перихондрію; потім до нього приєднується внутрішнє окостеніння хряща - ендохондральне окостеніння; тільки пізніше, коли в діафізах з'являються шляхом розпаду кістки порожнини кісткового мозку, епіфізи починають окостенівати ендохондрально; між діафізою і епіфізами, однак, довго залишається хрящ, за допомогою якого здійснюється зростання кістки в довжину з збереженням пропорцій (рис. 530).
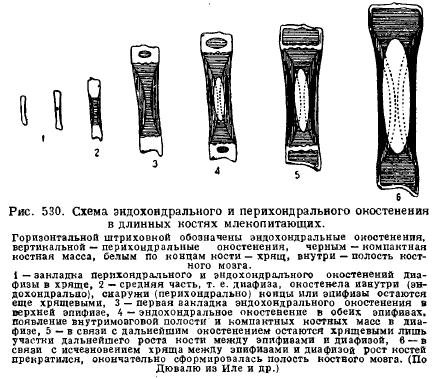
Хребетссавців характеризується на відміну від Sauropsida відсутністю вільних зчленувань між хребцями, за винятком першого та другого хребців. Між тілами хребців залягають міжхребцеві еластичні сполучнотканинні диски. Загалом це обумовлює меншу гнучкість хребта, що стоїть у зв'язку з більш спеціалізованим рухом ссавців за допомогою кінцівок.
Окостеніння тіл хребців відбувається за допомогою хрящових епіфіз. Тіла мають або плоскі поверхні або слабо опуклі, у більшості сучасних копитних злегка опістоцільні. На хребцях добре розвинені, особливо у грудній ділянці, остисті відростки, а також поперечні відростки, що сидять на дугах; у підстави останніх є сочленовні відростки.

Число шийних хребців, як правило, 7 (рис. 531), незалежно від довжини шиї, так що довжина шиї, на противагу птахам, залежить виключно від довжини хребців. Тільки у ламантину (Manatus) і в одного виду лінивця (Choloepus hoffmani) 6 шийних хребців, тоді як у трипалих лінивців (Bradypus) їх 9. Майже зникла у зв'язку з водним способом життя шия китоподібних все ж таки містить 7 плоских, частково зливаються хребців, тоді як у шиї жирафа ці ж 7 хребців дуже довгі. Перші 2 шийних хребця видозмінені в атлас і епістрофей, з'єднані справжніми суглобами, причому у однопрохідних зубоподібний відросток функціонує ще у вигляді особливої кістки, а не зрісся, як у інших ссавців, з тілом епістрофея. Рудиментарні ребра шийної області зростаються з хребцями, унаслідок чого утворюються продірявлені поперечні відростки. Часткове або повне зрощення шийних хребців має місце у китоподібних; у тушканчиків зростаються всі шийні хребці крім 7-го, у сумчастого крота (Notoryctes) зливаються від 2-го до 6-го, у крота - від 3-го до 4-го.
Число грудних хребців, які характеризуються причленуванням до них ребер, варіює від 9 [у одного китоподібного (Нyperoodon) та броненосця (Tatusia)] до 25 [у лінивця], але зазвичай їх буває 12-13. На грудних хребцях добре розвинені відростки: остисті (proc. spinosus), поперечні, або діапофізи (proc. transversus), сочленовні, або зигапофізи (proc. articularis); ребра причленовуються головками до тіла одного або двох сусідніх хребців та горбками до поперечних відростків. Вони складаються з двох частин: кістяного ребра – проксимального та хрящового – дистального. Тільки ребра передніх грудних хребців сягають грудини, задні ребра закінчуються вільно (хибні ребра). Поперекових хребців, які характеризуються відсутністю ребер, що увійшли до складу поперечних відростків, у ссавців від 2 до 9, зазвичай їх 6-7.
Хрестові хребці ссавців - це ті хребці, які, зрощені між собою, зчленовуються з клубової кісткою. Таких хребців 1-10, але тільки 1-2 з них є істинними крижовими хребцями, тобто такими, у яких зчленування з тазом відбувається за допомогою самостійних окостенінь, що відповідають крижовим ребрам; решта крижових хребців є помилковими, тобто прирослими до крижів хвостовими хребцями.
Число хребців, що входять в хвостовий відділ, варіює надзвичайно: від 3 ( летючі миші, Гіббон) до 49 [ящір (Manis macrura)].
Грудина(sternum) ссавців розвивається зі злиття черевних кінців ребер, які потім утворюють одну непарну пластинку, що складається з низки відділів (рис. 532).
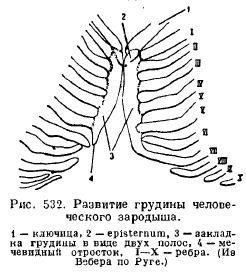
З'єднаний з ключицями (якщо вони є) і з першою парою ребер розширений передній відділ грудини зветься рукоятки (praesternum, manubrium); далі слід відділ, званий тілом (mesosternum, corpus), нерідко розпадається на ряд кісточок, послідовно розташованих між наступними парами ребер; Ще далі лежить так званий мечовидний відросток (xiphisternum). Не всі ребра перебувають у зв'язку з грудиною, більш-менш значне число помилкових ребер закінчується вільно (рис. 533).

У клоочних ссавців (Monotremata), у зв'язку з наявністю відсутніх у всіх інших ссавців коракоїдів, попереду від рукоятки грудини закладається хрящ передгрудинник (prosternum), що знаходиться у зв'язку з коракоїдами, який потім витісняється кісткою, що розвивається, рептилії.
Черепссавців цілу низку особливостей відрізняється від черепа рептилій. Однак ці особливості з'явилися не відразу, а шляхом поступової зміни, з якою можна познайомитися, вивчаючи черепи копалин звіроящерів (Theromorpha), особливо з загону звірозубих (Theriodontia). Череп ссавців – череп тропібазальний, але з укороченою міжочковою частиною, завдяки чому він виглядає як платибазальний, з великою черепною коробкою, з ясно вираженою тенденцією до зрощення окремих кісток між собою,
Про розташування кісток у черепі ссавців можна судити за схемою, що додається (рис. 534).
На бічних потиличних кістках, назовні від сочленовних горбків, зазвичай є потиличний відросток (рос. paroccipitalis), особливо добре розвинений у гризунів та у багатьох копитних. Попереду від сочленовних горбків зазвичай є отвір для виходу назовні під'язиковий (XII) нерв (for. condyloideum anterius).
У хрящі слухової капсули закладається кілька окостенінь, які, однак, дуже рано зливаються в дві кістки: попереду лежачу - кам'янисту (petrosum) і зовні і ззаду - соскоподібну (mastoideum); обидві ці кістки, зростаючись разом, дають кам'янисто-соскоподібну, або просто кам'янисту кістку (petromastoideum, petrosum або perioticum), що містить внутрішній слуховий лабіринт. Perioticum межує в задній частині з основною та бічною потиличними кістками, а в передній частині з клиноподібною та крило-клиноподібними кістками; Оскільки між цими кістками і кам'янистою кісткою проходять судини і нерви, то тут залишаються значні щілини: ззаду - задній рваний отвір (for. lacerum posterius), або яремний отвір (for. jugulare) для проходження язикоглоткового (IX) нерва, гілок блукаючого ( X) нерва і для додаткового (XI) нерва, а також для внутрішньої яремної вени, а спереду - передній рваний отвір (for. lacerum anterius), через який проходить у череп внутрішня сонна артерія. Іноді зв'язок кам'янистої кістки з черепом настільки слабка, що вона легко може бути відділена разом з барабанною кісткою, що приросла до неї, або навіть при мацерації черепа абсолютно випадає, як це має місце у китоподібних.
Зовні вушної області утворюються дві покривні кістки. Одна з них, походження якої до цих пір залишається ще мало з'ясованим, розвивається в області зовнішнього слухового проходу у вигляді півкола невеликої кісточки. Це – барабанна кістка (tympanicum), яка, ймовірно, є гомологом кутової кістки (angulare) нижньої щелепирептилій, що перейшла у привушну область разом з іншими кістками нижньої щелепи у зв'язку зі змінами, що відбулися у щелепному зчленуванні. Розташовуючись спочатку у вигляді півкола або кільцевої кістки в області слухового проходу, вона нерідко розростається і утворює, зазвичай разом з кам'янистою кісткою і іноді з лускатою, зовнішній слуховий прохід (meatus auditorius externus). На барабанній кістці буває натягнута барабанна перетинка. Глибша всередину барабанна кістка може брати участь також у освіті стін середнього вуха. Нерідко барабанна кістка на нижній стороні слухової області здувається в тонкостінний міхур (bulla ossea).
Друга кістка шкірного походження, що бере участь в утворенні кістяка вушної області, - це лускаста кістка (squamosum); вона отримала у ссавців винятковий розвиток та значення у зв'язку із причленуванням до неї нижньої щелепи. У мавп та людини всі ці кістки вушної області зростаються разом, утворюючи складну скроневу кістку (temporale); луската кістка тут становить лише частину, саме луску (squama temporalis) скроневої кістки, тоді як барабанна кістка - барабанну частину (pars tympanicum), а власне привушні хрящові кістки утворюють частини кам'янисту (pars petrosa) та соскоподібну (pars mastoidea). Ступінь розвитку лускатої кістки та участь її в утворенні стінок мозкової коробки та слухового проходу дуже різні, але завжди луската кістка має счленовную поверхню (саvitas glenoidea) для причленування нижньої щелепи. Таким чином, у ссавців на відміну від решти хребетних є абсолютно особливе оригінальне причленування нижньої щелепи: не до квадратної кістки, а до лускатої кістки черепа. Счленовна поверхня для нижньої щелепи лежить на підставі того, що йде від луски назовні і вперед вилицевого відростка(proc. zygomaticus). У однопрохідних над сочленовною поверхнею він пронизаний скроневим каналом (can. temporalis), особливо добре вираженим у качконоса. Позаду сочленівної ямки є відросток (рос. postglenoideus).
Спереду від слухового відділу основа черепа утворена основною клиноподібною кісткою (basisphenoidale), на внутрішньої поверхніякої є більш менш значне поглиблення - так зване турецьке сідло (sella turtica), в якому міститься гіпофіза. З боків від основної клиноподібної кістки лежать, утворюючи нижні частини передньої стінки черепної коробки, крило-клиноподібні кістки (alisphenoidea), які зазвичай зростаються з нею в одну кістку, утворюючи великі крила (alae magnae) цієї кістки. Крило-клиноподібні кістки забезпечені двома отворами: овальним (for. ovale) для проходження третьої гілки трійчастого нерва (V3) та круглим (for. rotundum) для проходження другої гілки того ж нерва (V2). Втім, перше нерідко зливається з переднім рваним отвором (for. lacerum anteris), а друге часто зливається з клино-очковим отвором (for. sphenorbitale), обмеженим одночасно як крило-клиноподібною кісткою, так і лежачою попереду від неї очно-клиновидною. Через цей отвір проходять нерви око-двітельний (oculomotorius III), блоковидний (trochlearis IV), що відводить (abducens VI) і перша гілка трійчастого нерва.
Черепна кришка тут прикрита тім'яними кістками (parietale), які нерідко зростаються один з одним (у однопрохідних, деяких сумчастих і копитних); між ними і верхньою потиличною кісткою закладаються ще дві кістки, що зливаються в одну міжтем'яну кістку (interparietale), яка в деяких загонах зберігається у вигляді самостійної кістки (китоподібні, сумчасті, дамани, трубкозуб, шерстокрил і деякі гризуни), тоді як в інших вона зростається або з тім'яними (у деяких гризунів, сирен і жуйних), плі з верхньою потиличною кісткою (у хижих, однокопитних, приматів).
Попереду основної клиноподібної кістки лежить передня клиноподібна кістка (praesphenoideum) із заглибленням для хіазми очних нервів; з боків від неї, зазвичай зростаючись з нею, лежать очно-клиноподібні кістки (orbitosphenoidea) з отвором для проходження очних нервів (for. opticum), звані у разі зрощення з передньою і основною клиноподібними кістками малими крилами (alae parvae). Як уже сказано вище, між обома крилоподібними кістками з кожного боку є щілинні отвори, через які проходять нерви очних м'язів і перша гілка трійчастого нерва (for. sphenorbitale).
Зверху цьому ряду кісток відповідають покривні лобові кістки (frontalе) - зазвичай добре розвинені парні кістки, що іноді зростаються між собою і несуть у положніх стрижні, а у оленів - пеньки для рогів. Покриваючи черепну коробку, лобові кістки обмежують зверху і з внутрішньої сторони очниці, утворюючи нерідко відростки: передочковий (роз. praeorbitalis і поза-очний (роз. postorbitalis). Іноді навколо очей утворюється внаслідок зрощення вилицевої кістки(jugale) з лобовою кісткою навколоочкове кільце (деякі комахоїдні та хижі, жуйні, однокопитні, напівмавпи). У мавп процес роз'єднання очної западини від скроневої йде так далеко, що між очницею та скроневою западиною утворюється кістяна стінка.
У передній стінці очниці лежить невелика покривна слізна кісточка (lacrymale), продірявлена отвором слізного каналу (for. lacrymale).
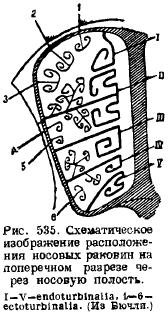
У носовій ділянці закладається в хрящовій носовій капсулі кілька окостенінь. По-перше, вздовж хрящової носової перегородки розвивається посередині кісткова перегородка (lamina perpendicularis), що відповідає середній нюхової кістки (mesethmoideum); по-друге, в задній стінціносової капсули і в передній стінці мозкової порожнини розвиваються дві бічні пластинки, продірявлені численними отворами для проходження нюхового нерва - решітчасті пластинки (lamina cribrosa), відповідні бічним етe . Від носової перегородки окостеніння поширюється з дорзального боку з обох боків, йде з бокових стінках носових капсул і закінчується на дні, утворюючи зі сторін численні виступи всередину носової порожнини. Це нюхові раковини (ethmoturbinalia), що утворюють разом із бічними пластинками і з порожнинами комірчастої будови іноді дуже складний носовий лабіринт. Розташування раковин видно з схематичного малюнка, що додається (рис. 535). Від покривної носової кістки (nasale), що покриває зверху цю область, звисають усередину носової порожнини, теж з одного боку, раковини (nasoturbmalia). Знизу на носову перегородкуналягає непарна кістка, що вважалася раніше сошника (vomer). Однак дослідження останнього часу показали, що, крім цієї непарної покривної кістки, в основі черепа розвиваються дві парні кістки, названі передсошником (praevomer); а так як у рептилій сошник є у вигляді парної кістки, а непарною кісткою в цій області був парасфеноїд, то гомологом цієї останньої кістки рептилій можна вважати саме сошник ссавців, а передсошники вважати сошниками. Спереду стінками носової порожнини є щелепні кістки (maxillare), у яких утворюються раковини, вростають у носову порожнину; це - щелепні раковини (maxilloturbinale).
Лицьова сторона черепа закінчується спереду міжщелепними кістками (praemaxillare). Ця кістка в антропоморфних мавп і в людини дуже рано зростається з щелепною кісткою, а у рукокрилих редукована до невеликої кісточки або навіть зовсім відсутня. Своїм вертикальним відростком міжщелепна кістка обмежує спереду носовий отвір і заходить між носовою кісткою та щелепною далеко назад, іноді до лобової кістки. Знизу міжщелепні кістки беруть участь в утворенні твердого піднебіння. Вони обмежують спереду і з боків на твердому небі отвори піднебінного (стенсонового) каналу (for. incisiva), що з'єднує ротову порожнинуз якобсоновим органом. Що лежать назовні цих піднебінно-носових отворів відростки міжщелепних кісток накладаються для кожної кістки незалежно, ймовірно, є гомологами парного сошника рептилій (praevomer) і у качконоса зберігаються у вигляді самостійних кісток. На міжщелепній кістці розташовуються різці.
Щелепні кістки (maxillare), як було зазначено, обмежують дно носової порожнини, утворюючи тверде піднебіння горизонтальними відростками. Про внутрішню сторону носової порожнини до бічних стінок її, утворених щелепними кістками, приростає нижня раковина (maxilloturbinae), що носиться, що закладається як самостійна кістка.
Продовженням твердого піднебіння тому служать горизонтальні відростки піднебінних кісток (palatinum), що обмежують як бічні стінки внутрішній носовий прохід. Вони стоять вертикально, і до них ззаду примикають також крилоподібні кісткою (pterygoideum), що вертикально стоять, у людини зростаються з клиноподібною кісткою, утворюючи її крилоподібні відростки (proc. pterygoidei).
Значні видозміни зазнає переднього відділу черепа у зв'язку з переходом до водного способу життя у китоподібних, сирен та у зв'язку з розвитком хобота у слонів.

У китоподібних (мал. 536) ніздрі відсунуті далеко назад у міжочкову область і ведуть у вертикальний канал, що відповідно відкривається хоанами в ротову порожнину; носові кістки лежать за ніздрями, налягаючи на лобові кістки, і майже прилягають до межтеменной кістки; також і щелепні кістки налягають на лобові кістки, заходячи майже до тім'яних кісток, сильно зрушених на боки міжтемної кістки.
У сирен має місце таке ж переміщення назад ніздрі, але не так далеко; носові кістки тут ще більше редуковані і навіть можуть бути відсутніми (рис. 537).

У слонів з непомірним розвитком бивнів та потужних корінних зубів та у зв'язку з наявністю хобота сильно видозмінюється нормальне розташуваннякісток: ніздрі сильно відсунуті назад, у зв'язку з цим носовий канал набуває майже перпендикулярного положення; лобові кістки дуже короткі, міжщелепні кістки дуже великі та заходять далеко назад; щелепні кістки зсунуті під основу черепа; весь череп робиться дуже масивним із надзвичайно розвиненою пневматичністю кісток (рис. 538).

Нижня щелепа ссавців на відміну від решти хребетних розвивається з кожного боку тільки з однієї зубної кістки (dentale); обидві половини або залишаються з'єднаними один з одним за допомогою хряща, шва, плі ж зростаються між собою (однокопитні, слон, рукокрилі, мавпи). Кожна половина складається з горизонтального тіла (corpus), на якому сидять зуби, і висхідної частини з відростками: переднім вінцевим відростком (рос. coronoideus), що лежить позаду нього счленовним відростком (рос. articularis) і у нижчих ссавців кутовим відростком, що йде назад angularis); у більшості сумчастих і в окремих гризунів цей відросток перетворюється на пластинку, що простягається всередину (рис. 539).
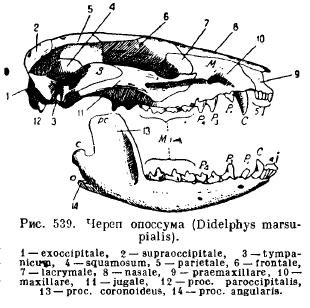
Така будова нижньої щелепи і своєрідне її приєднання до черепа приводять нас до питання щодо походження цих особливостей у ссавців. Історія індивідуального розвиткуі палеонтологічні дані дають у відповідь те питання. У ембріонів ссавців закладаються піднебінно-квадратний та мекелевий хрящі. Однак окостеніння піднебінно-квадратного хряща відбувається поза зв'язком із зчленуванням нижньої щелепи і не призводить до утворення квадратної кістки, як у рептилій, і дасть нову слухову кісточку - ковадло (incus), тоді як мекелевий хрящ, окостеневий у своїй задній частині, не дає. кістка (articulare), як у інших щелепноротних, а третю слухову кісточку - молоточок (malleus). Перша слухова кісточка, що прагне (stapes), відповідає стовпчику амфібій, рептилій і птахів і, як ми вже знаємо, є видозміненою підвіскою (hyomandibulare), тобто проксимальним відділом під'язичної дуги. Так само як і стовпчик (columella auris), стрем'я прикриває овальне вікно слухового лабіринту, і продірявлено, як у деяких рептилій, отвором, через який у деяких нижчих ссавців проходить лицьова артерія (а. facialis) (рис. 540 і 54).

Таким чином, у ссавців квадратна кістка перетворилася на ковадло, сочленівна - на молоточок; бо до молоточку приростає ще невелика покривна кісточка, що утворює середній його відросток, то вважають, що це останній відповідає покривної сочленовной кістки (goniale) рептилій. Покривна кутова кістка рептилій, що лежить по сусідству з цією останньою кісточкою, також переходить у вушну область, стаючи барабанною кісткою (tympanicum). Зазначені зміни видно на схемі, що додається (рис. 541).
Палеонтологічні дані підтверджують можливість уканених вище перетворень. Серед звірозубих звіроящерів (Theriodontia) зустрічаються такі, у яких зубна кістка набуває особливо великого розвитку, досягаючи ззаду місця зчленування нижньої щелепи з квадратним пензлем. У той же час луската кістка спускається квадратною кісткою до цього ж зчленування. Зубна і луската кістки, таким чином, дедалі більше зближуються.
Нарешті, у деяких звірозубих зубна і луската кістки беруть участь у зчленуванні поруч із сочленовною та квадратною кістками.
Що стосується під'язичного апарату ссавців, то він складається з тіла, видозміненої базальної частини (copula) гіоїдної дуги, з передніх ріжків (cornua anteriora), утворених нижнім відділом гіоїдної дуги, і задніх ріжків (cornua posteriora), перетворених з першої зябрової. Передні ріжки складаються з кількох окостенінь (до чотирьох); звернений до черепа відрізок часто приростає до кам'янистої кістки, утворюючи шиловидний відросток (proc. styloideus).
Друга та третя зяброві дуги дали початок утворенню щитовидного хряща (cartilago thyreoidea.)
Пояси кінцівок.Плечовий пояс ссавців характеризується насамперед редукцією воронових кісток (procoracoideum, epicoracoideum, coracoideum), тенденцією до зникнення ключиць (clavicula) та добре розвиненим гребенем лопатки (spina scapulae), що продовжується в акроміальний відросток (acromion).
Однак у однопрохідних плечовий пояс має велику подібність до рептилій. Лопатка має акроміальний виступ, до якого прилягає спереду ключиця, а ззаду до лопатки причленовуються широкі і короткі воронові кістки (coracoideum), що йдуть до грудини. Спереду від них є у вигляді самостійних кісток широкі прокоракоїди (procoracoideum). Ключиці (clavicula) медіально сполучаються з надгрудником (episternum) (рис. 542). У інших ссавців кракоїди закладаються у вигляді двох окостенінь, але втрачають зв'язок із грудиною і приростають до лопатки, утворюючи коракоїдний відросток (proc. coracoideus); прокоракоїди зникають зовсім (рис. 543), ключиці зникають у деяких хижих і копитних у зв'язку з пристосуванням до швидкого бігу, а у китоподібних і сирен у зв'язку з перетворенням передніх кінцівок на плавники.

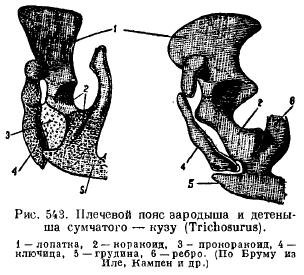
Тазовий пояс ссавців має при закладці типове розташування елементів; особливістю його є, по-перше, наявність додаткової кістки у вертлужній ямці (acetabulare), по-друге, зрощення всіх цих кісток з кожної сторони в одну безіменну кістку (innominatum) і зрощення між собою лобкової та сідничної кісток, так що утворюється замкнуте з всіх сторін отвір між цими кістками (for. obturatum) (рис. 544). У однопрохідних і сумчастих до лобкової кістки знизу причленовується особлива кістка підтримки сумки (marsupiale).
Скелет кінцівоку вихідних форм ссавців зумовлений їх пристосуванням до швидких рухів - бігу за допомогою чотирьох кінцівок, що розташовуються під тулубом, так що останнє не висить між кінцівками, як у більшості рептилій, а безпосередньо на них спирається. Здатність до бігу стоїть у зв'язку з іншим становищем кінцівок у ссавців. Саме в них кінцівки орієнтовані так, що ліктьові суглоби передніх кінцівок направлені назад, а колінні зчленування задніх – уперед. При рухах передні кінцівки викидаються вперед і притягують тулуб згинанням кистового і ліктьового суглобів; задні кінцівки відштовхують тулуб уперед шляхом випрямлення ноги в колінному суглобі.
За кількістю вхідних до складу кінцівок кісток і за їх розташуванням скелет кінцівок родоначальних форм ссавців має дуже типову будову примітивної п'ятипалої кінцівки і піддався великим змінамв межах класу лише в процесі пристосування до різноманітних рухів у різних умовахсередовища.
У передній кінцівці характерна наявність у дистальному відділі плечовий кісткиотвори (for. entepicondylare), наявність проксимального виступу ліктьової кістки (olecranon), наявність горохової кісточки (pisiforme) і злиття четвертої та п'ятої дистальних карпальних кісточок (carpalia dis talia 4+6 = hamatum, або uncinatum) і центральної проксимальної dist. + intermedium = lunatum, або semilunare). У зв'язку з тим, що часто використовується номенклатура кісточок зап'ястя ссавців, прийнята в анатомії людини, наводимо таблицю цих назв у схемі, що вказує на розташування кісток.
Задня кінцівка ссавців характеризується насамперед тим, що повернена колінним зчленуванням попереду стегно має добре розвинені на проксимальній частині відростки - вертлуги в числі 2-3 (trochanter major, minor та tertius); в гомілки великогомілкова кістка розвинена сильніше, є кісткова колінна чашечка (patella), що захищає колінний суглоб. У передплюсні є тільки дві проксимальні кістки, добре розвинені: внутрішня - таранна (talus = astragalus), що утворилася від злиття проміжної та центральної проксимальної (intermedium + centrale proximale), і зовнішня п'яткова (calcaneus), що відповідає фібулярній кістці. З кісточок середнього ряду центральна дистальна (centrale distale) перетворилася на човноподібну (naviculare = scaphoideum). Тибіальна кісточка або редукувалась або приросла до цієї останньої.
Схема розташування кісточок передплюсни та плюсни та синоніміка назв такі:
У зв'язку з пристосуванням до пересування в різних середовищах кінцівки ссавців зазнали дуже суттєвих змін, причому ці зміни особливо добре можна простежити на скелеті. На жаль, палеонтологічних даних про будову кінцівок родоначальників ссавців ми не маємо. Безсумнівно, що з перших ссавців мало спеціалізовані кінцівки описаного вище типу. Такі саме кінцівки мають із сучасних нам ссавців найпримітивніші сумчасті, наприклад, опосуми - сумчасті щури (Didelphys), і комахоїдні, що ведуть деревний спосіб життя. Проте було б неправильно робити звідси висновок, що родоначальниками ссавців неодмінно були жителі лісів і до того ж лазали по деревах форми. До того ж кінцівка опосума, хоч і служить для пересування по сучках дерев, є аж ніяк не типовою спеціалізованою дерев'яною кінцівкою, а зберігає свій універсальний тип. Такі кінцівки могли найкраще зберегтися у тварин, не тільки бігали по землі, а й підіймалися на дерева, а також у тварин, що лазили горами. Спеціалізація кінцівок ссавців йшла одночасно з диференціацією загонів та інших дрібніших систематичних одиниць, причому мали місце конвергенції.
Особливо глибоким видозмінам піддаються кінцівки при пристосування до польоту, до плавання у воді, до пересування під землею; повільні рухи по землі та деревний спосіб життя не стоять у зв'язку з глибокою спеціалізацією.
У кажанів (Chiroptera) передня кінцівка перетворена на крило, причому літальна перетинка натягується між чотирма (2-б) сильно подовженими пальцями, позбавленими пазурів; перший палець короткий і має на кінці сильно розвинений кіготь; цей палець не грає жодної ролі при літанні, служить для лазіння; у зап'ясті човноподібна, півмісячна та центральна кісточки зростаються в одну; передпліччя подовжене і складається з добре розвиненої променевої кістки, тоді як ліктьова кістка редукована і приростає до променевої. Літальне перетинання захоплює і задні ноги. Між останніми вона розвинена у рукокрилих по-різному (рис. 545).
У плаваючих у воді ссавців кінцівки перетворюються на плавники так само, як це ми вже бачили у рептилій. Особливо сильно змінилися при цьому кінцівки у китів та сирен. На противагу крилу передня кінцівка значно коротшає у своїй проксимальній частині. У китів кістки плеча і передпліччя перетворилися на короткі та широкі платівки, що дають місце для прикріплення потужної мускулатури. Усі відділи кінцівки нерухомо з'єднані один з одним, утворюючи міцне весло. У цьому число фаланг може бути збільшено. Те ж зміни ми бачили у рептилій. Збільшення числа фаланг залежить від того, що діафізи фаланг не зростаються з епіфізами і окостеніють з окремих центрів. Таке окреме окостеніння стоїть у зв'язку з уповільненим процесом окостеніння цих тварин. Уповільненість процесу окостеніння веде до того, більшість кісток у місцях з'єднання їх залишається хрящовими.
Плавники сирен (Sirenia) мають з такими китоподібними лити зовнішню подібність: плече і передпліччя між собою зчленовані, так що можуть згинатися; є майже всі кісточки зап'ястя і п'ять пальців з малозміненим числом фаланг (рис. 546).
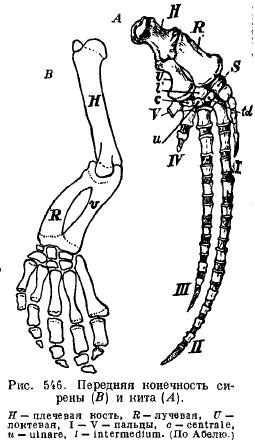
Задні кінцівки у сучасних китіві сирен редуковані настільки, що від них залишаються лише залишки тазу (рис. 547 та 548).
Інший тип пристосування кінцівок до пересування в поді бачимо у ластоногих (Pinnipedia). Тут хвіст майже зникає, а задні ноги пересуваються назад, граючи роль рухового органу. Три сімейства цього загону представляють за рівнем розвитку пристосованості до пересування у воді хіба що три щаблі (не генетично). У моржів і у вухатих тюленів (Otariidae) задні кінцівки ще можуть підгинатися вперед під тулуб, але у моржів у пристрої кінцівок більше примітивності, вони більш короткі та рухливі. У тюленів (Phocidae) задні ноги вже не можуть підгинатися під тіло вперед, проте на пальцях ніг ще зберігаються пазурі.
У тварин, що риють, кінцівки піддаються також дуже сильним видозмінам: особливо сильно змінилися кінцівки сумчастого крота (Notoryctes typhlops). У нього на лопатці є два гребені, як і у багатьох тварин, що риють, що стоїть у зв'язку з сильним розвитком триголового м'яза плеча; є ключиця, як у всіх ссавців, що риють. На плечовій кістці є дуже сильний гребінь, що розгалужується вилкоподібно і служить для прикріплення сильних (необхідних при копанні) дельтовидної і грудного м'яза; сильно розвинений внутрішній виросток плеча завдяки сильному розвиткум'язів згиначів ліктьової кістки (proc. olecranon), який служить місцем прикріплення триголового м'яза; зап'ястя своєрідно змінено, стаючи дуже міцним; 1-й і 2-й пальці відсунуті на долонний бік, і для копання служать лише 3-й і 4-й, забезпечені величезними кігтями, особливо 3-й; число фаланг у пальців дуже мало: 2 і 1. 5-е метакарпале з широким плоским пазуром, що збільшує риючу поверхню; в площині долоні знаходиться сесамоподібна кісточка, до якої прикріплюється м'яз, що згинає пальці, що грає велику роль при копанні; це збільшує міцність зап'ястя; як і в інших риючих форм, симфіз лобкових кістокдуже короткий; сіднична кістка зростається з поперечними відростками обох останніх крижових хребців; на стегні, коліні та великогомілкової кістки сильно розвинені гребені та вирости - місця прикріплення м'язів; великогомілкова і малогомілкова кістки (tibia і fibula) міцно зростаються, як у златокрота (Chrysochloris); підошва відчинена назовні, як і у Chrysochloris; 5-й палець загнутий на підошовну поверхню, 5-й ніготь на ступні широкий і лопатоподібний, 4-й довгий і серповидний, 3-й і 2-й стають коротшими і ширшими; на грудній кістці сильно розвинений гребінь, як у златокрота, сліпуша (Spalax) та крота (Talpa) (рис. 549).
Сильне розвиток гребенів на кістках для прикріплення м'язів, укорочення carpus, міцне з'єднання carpalia, редукція одних фаланг і посилення інших, що грають роль при копанні, - всі ці ознаки властиві ссавцям, що риють взагалі. Але в залежності від того, в якому грунті тварина риється, будуть спостерігатися відмінності. При копанні в м'якому грунті (наприклад, у крота - Talpa) (рис. 550) кисть сильно розширюється, вона діє як лопата і має схильність до розвитку 6-го пальця. Передня кінцівка крота можна порівняти з лопатою, що служить для копання в м'якому грунті. Навпаки, у тих форм, які риються в твердому грунті, кисть стає вже, в копанні беруть участь лише деякі пальці, здебільшого 3-й і 4-й, що розвиваються сильніше за інші, як у Notoryctes і Chrysochloris. Ця кисть можна порівняти із заступом для копання в твердому грунті.
![]()
Те ж бачимо ми у ссавців, що розривають тверді будівлі термітів, наприклад, у мурахоїда (Myrmecophagus), у тамандуа (Tamandua). Ці тварини ходять, спираючись на зовнішній край ступнів, підігнувши пазурі.
Пристосування ссавців до швидкого пересування землею викликали дуже істотні зміни у кінцівках. Пересування по землі, яке було властиве примітивним ссавцям, здійснювалося шляхом опори на всю кисть та ступню; вихідні формиссавців були первинно стопоходящими або плантиградними, тобто спиралися при ходьбі на всю ступню, як, наприклад, ведмеді. Але така будова допускає лише відносно повільне ходіння в порівнянні з швидконогими антилопами або кіньми, або зайцями, що стрибають, кенгуру, тушканчиками. Протягом філогенетичного розвитку ми бачимо поступове підняття над землею зап'ястя і передплюсни (carpus і tarsus,) за якими потім пішли п'ясти і плюсни, але фаланги ще стосувалися землі цілком. Зрештою, випрямилися і фаланги, передні та задні кінцівки стали спиратися на кінці пальців. Етапи цієї еволюції ми можемо тепер простежити крок за кроком не тільки у копитних, а й у хижих ссавців, які набули здатності до швидкого бігу. У міру випрямлення кінцівок крайні пальці перестають стикатися із землею, перестають функціонувати та піддаються атрофії. Виходить менша поверхня зіткнення із землею. Зміни в дистальній частині кінцівок спричиняли зміни в проксимальній: carpalia і tarsalia розташовуються все більш щільно один біля одного, ulna і fibula редукуються, кінцівки починають здійснювати рухи лише в сагітальній площині, серединні metapodia зливаються в одну кістку.
Два шляхи розвитку ведуть до утворення кінцівок сучасних копитних. В одному ряду вісь кінцівки проходить у середині третього пальця і останній розвивається найсильніше, решта ж стають рудиментарними. Це - непарнокопитні (Perissodactyla), мезаксіальні (Mesaxonia). В інших вісь проходить між третім і четвертим пальцями, що розвиваються рівномірно: парнокопитні (Artiodactyla), параксіальні (Paraxonia). Бічні пальці редукуються. Розвиток непарнокопитних найнаочніше видно на філогенетичному розвитку кінцівок коня (рис. 551). Розвиток парнокопитного стану зумовлено було, між іншим, тим, що йому передувала редукція першого пальця, так що при переході від стоприходу він майже не брав ніякої участі, тоді як у однокопитних предків розвиток п'ятого пальця спочатку був нормальним,
Цікаво, що і парнокопитливість і непарнокопитливість виникали в еволюційному процесі неодноразово і в деяких випадках призводили до дуже спеціалізованого типу, наприклад, у американських Proterotlieriidae (вимерлих).
Пальці, що зберігаються, одягнені копитами, які можуть диференціюватися далі в залежності від способу життя і місцеперебування. Так, у козлів, що тримаються на скелях, копита круті та вузькі та забезпечені гострим краєм; у північного оленявони дуже довгі, широко роздвоєні і служать свого роду лижами, що не дають тварині провалюватися в сніг та болота.
Одночасно з пальцями при розвитку ходіння на пальцях, відбувається зміна зап'ястя (carpus) та передплюсни (tarsus). Воно йде по-різному у парнокопитних і непарнокопитних у зв'язку з іншим розподілом сили тяжіння. Нарешті, у зв'язку з переходом до нальцехождіння помалу зменшується значення ліктьової кістки (ulna) як опори для плеча: вона зменшується або зливається з променевою (radius). Так само редукується і, нарешті, зникає в задніх кінцівках малогомілкова кістка (fibula).
Значення та характер описаних змін ясно видно з порівняння кінцівки верблюда та слона (рис. 552). У слона кисть (стопа) залишається короткою, а подовжені плече та передпліччя (стегно та гомілка). Навпаки, у верблюда подовжуються зростаючі metapodia; плече та передпліччя (а також стегно та гомілка) залишаються короткими. Завдяки випаданню ряду кісток відбувається спрощення кістяка кінцівки, остання стає легшою. Редукція пальців веде до меншого тертя, зменшується небезпека ушкодження, розриву зв'язок. Зростання metapodia створює велику міцність. Кінцівка слона набагато важча. Велика кількість кісток у пензлі вимагає для отримання стійкості з'єднання всіх пальців. загальною шкірою. Кінцевість слона – це колона великої міцності, але позбавлена тієї легкості, яку має кінцівка верблюда.
Іншим способом швидкого пересування на землі є стрибання. Це пристосування виникало неодноразово в історії ссавців і до того ж у найрізноманітніших філогенетичних стовбурах: у ряді сумчастих – у кенгуру (Macropus), серед комахоїдних – у стрибунців (Macroscelidae), і, нарешті, у різних рядахрозвитку гризунів: сем. зайці (Leporidae) із Duplicidentata; у ряді дикобразоподібних (Hystricomorplia) розвинулася група довгоногів (Pedetinae); у ряді мишеподібних (Myomorpha) ми маємо австралійського тушканчика (Conilurus), тушканчиків Старого Світу (Jaculus, Alactaga), американських стрибунців (Zapus) та мішчастих стрибунців (Dipodomys).
Розвиток здатності до стрибання пов'язане з подовженням задньої кінцівки, зокрема кісток плюсни (metatarsus). Крім того, при цьому відбувається втрата зайвих крайніх пальців, і передплюсні кістки з'єднуються між собою дуже міцно. Прикладом можуть бути ноги земляного зайця (Alactaga) і реальних тушканчиків (Dipus).
Передні ноги у Alactaga малі та короткі. Земляний заєць користується ними при копанні, але не стосується ними землі при стрибанні. Стрибка здійснюється лише за допомогою задніх ніг і навіть без хвоста. Останній стосується землі лише за спокійному становищі, коли тварина спирається на хвіст. Скелет ноги має ту особливість, що плюсневі кістки 2-го, 3-го та 4-го пальців зливаються в одну кістку. Кінці плеснових кісток унизу не злиті, і серединна (3-й палець) виступає далі вперед. Зі сполучених з ними фаланг знов-таки середні найдовші. 1-й та 5-й пальці рудиментарні і не стосуються землі. Обидва ці пальці мають однакову довжинузавдяки тому, що плюснева кістка великого пальцядовше, ніж 5-го. Перша має 2, друга – 3 фаланги.
У Dipus спеціалізація пішла ще далі: перший палець зник без сліду, від 5-го зберігається лише рудимент. Середній палецьвдвічі слабше, ніж два крайні, і лише трохи довші за них. Пальці паралельні. Середній палець редукується тому, що його функцію беруть на себе 2-й і 4-й, на кінці яких доводиться упор при сидінні та стрибанні. У подальшій еволюції третій палець повинен зникнути: ми маємо у Dipus незакінчений тип пристосування.