What is the thoracic spine of mammals made of? The external structure of mammals, the skeleton and muscles on the example of a domestic dog
The spine consists of the cervical, thoracic, lumbar, sacral and caudal regions. His salient feature- platycelial (with flat surfaces) form of the vertebrae, between which are cartilaginous intervertebral discs. The upper arches are well defined. There are seven vertebrae in the cervical region, the length of which determines the length of the neck; only the manatee and the sloth - Choloepus hof-fmani have 6 of them, and the sloth - Bradypus 8-10. The neck vertebrae are very long in giraffes and very short in cetaceans, which do not have a cervical interception. The ribs that form the chest are attached to the vertebrae of the thoracic region. The sternum closing it is flat and only in bats and burrowing species with powerful forelimbs (for example, moles) has a small crest (keel), which serves as an attachment point for the pectoral muscles. In the thoracic region there are 9-24 (usually 12-15) vertebrae, the last 2-5 thoracic vertebrae bear false ribs that do not reach the sternum. In the lumbar region from 2 to 9 vertebrae; rudimentary ribs merge with their large transverse processes. The sacral region is formed by 4-10 fused vertebrae, of which only the first two are truly sacral, and the rest are caudal. The number of free tail vertebrae ranges from 3 (in the gibbon) to 49 in the long-tailed pangolin.
The degree of mobility of individual vertebrae is different. In small running and climbing animals, it is large, along the entire length of the spine, so their body can bend into different directions and even curl up into a ball. The thoracic and lumbar vertebrae are less mobile in large, rapidly moving animals. In mammals that move hind legs ax (kangaroos, jerboas, jumpers), the largest vertebrae are located at the base of the tail and sacrum, and further forward their size consistently decreases. In ungulates, on the contrary, the vertebrae and especially their spinous processes are larger in the anterior part of the thoracic region, where powerful muscles of the neck and partly of the forelimbs are attached to them (
The external structure of the dog
The dog was the first pet. Man tamed it in ancient times. The dog helped the primitive man during the hunt, guarded his dwelling. Now service, hunting and decorative dogs, as well as mongrels (outbred) are known. Dogs help to serve to protect the borders of our Motherland, and during the Great Patriotic War searched for the wounded, helped in reconnaissance and communications. On dogs, scientists conduct research on the vital activity of the organism. Dog before man visited space.
The body of a dog, such as a shepherd dog or a husky, is slender and muscular. Its limbs are not located on the sides of the body, as in reptiles, but under the body, so the body of the animal does not touch the ground. The dog walks, leaning on fingers with strong claws. The flexible neck allows for great head mobility. The mouth of the animal is limited by movable lips - upper and lower. Above upper lip there is a nose with a pair of external nasal openings - nostrils. The eyes have well developed eyelids. The nictitating membrane (third eyelid) in a dog, like in all mammals, is underdeveloped. Of all animals, only mammals have outer ear - Auricle. The outer ear of the dog is large and mobile.
Integuments of mammals
The skin of mammals is strong and elastic; in most animals, the bases of the hair are placed in it, which form the characteristic for this class of vertebrates. hairline . Distinguish between thick and long hair — awn and shorter, softer - underfur, or undercoat. Rough and strong awn protects the undercoat and skin from damage. The undercoat, which traps a lot of air, retains body heat well. In addition to the awn and underfur, animals develop big hair- organs of touch.
Mammalian hair, like bird feathers and reptile scales, is composed of horny matter. Twice a year, a dog, like many other mammals, sheds - part of its hair falls out and is replaced by new ones. The base of the hair is located inside a special hair bag, where the ducts of those nearby open. sebaceous glands. Their secretions lubricate the skin and hair, which makes them elastic and not wetted by water.
Figure: The structure of the skin and hair of a mammal
In the skin of most animals are located and sweat glands . Sweat, evaporating from the surface of the body, cools it. Together with sweat, excess salt and urea are also excreted from the body; thus, sweat glands play a role additional bodies selection.
At the dog sweat glands little and cooling of the body is achieved with increased breathing.
At the ends of the fingers of mammals are horny claws, nails or hooves. Sometimes horny formations are also developed on the head (horns in rhinos, antelopes, cattle, etc.) or on the tail (for example, horny scales in rats).
mammalian skeleton
The skeleton of mammals consists of the same sections as those of other vertebrates. The skull of animals is distinguished by a larger cranium, which is associated with large sizes brain. For mammals, the presence of 7 cervical vertebrae is very characteristic. Both the long-necked giraffe and the whales have the same number of cervical vertebrae. The thoracic vertebrae (usually 12-15) together with the ribs and sternum form a strong chest. massive vertebrae lumbar movably articulated with each other. In this section, the body can bend and unbend. The number of lumbar vertebrae varies in different types(2-9), the dog has 6 of them. The sacral spine (3-4 vertebrae) fuses with the pelvic bones. The number of vertebrae in the caudal region (from three to several dozen) depends on the length of the tail.

Figure: Mammalian skeleton on the example of a dog

Pattern: Dog skull
Forelimb belt mammals consists of two shoulder blades with crow bones attached to them and two clavicles. The dog's clavicles are not developed. Rear limb belt- pelvis - formed by three pairs pelvic bones. The skeletons of the limbs in different species of mammals and reptiles are similar, but the details of their structure in different species are different and depend on the living conditions of the animal.
Mammal muscles
In most mammals, including the dog, the muscles of the back, limbs and their belts are especially developed. The dog can run in large jumps, bending and straightening the body, alternately pushing off the ground with either front or hind legs. Strong muscles move the lower jaw. Clenching the jaws, armed with teeth, the dog holds the prey firmly.

Figure: Muscles of a dog that move the lower jaw
Shoulder girdle mammals is simplified, consists of a large shoulder blades, which has a coracoid outgrowth, and clavicle(Fig. 80, 81). At the embryonic stage of development and in adult monotremes coracoid is a separate bone.
shoulder blade located on top chest, has a triangular shape and a high outgrowth (comb) with acromial a process for attaching muscles. At the location of the coracoid process and the acromial process of the scapula is articular fossa for articulation of the forelimb.
Collarbone rod-shaped serves to connect the shoulder and sternum. The clavicle is most developed in animals performing with the forelimbs complex movements(bats, moles, primates, rodents, etc.). In many species that are characterized by monotonous movements (hoofed, canine), the clavicle is absent. In general, the forelimb girdle is connected to the axial skeleton through muscles and ligaments.
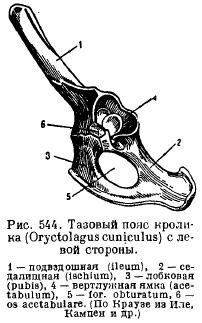
Pelvic girdle (Fig. 82 A) consists of paired nameless bones, formed by articulation iliac, ischial and pubic bones. Taz closed type and through the iliac bones is firmly connected to sacral department spine. When the elements of the pelvis grow together, pubic fusion and formed acetabulum for articulation of the hind limb. In the lower part of the innominate bone there is obturator foramen.
Skeleton of paired limbs
The limbs of mammals, like those of other terrestrial vertebrates, are five-fingered and represent a three-membered lever, all parts of which are movably connected to each other.
forelimb consists of shoulders, forearms and hands(Fig. 81). Brachial bone well developed, has a rounded head for attachment to the limb girdle. Forearm make up two bones radial and ulnar, which has olecranon at the top. At the junction of the shoulder and forearm is elbow joint.
Brush includes three departments - wrist, metacarpus and phalanges of fingers. Wrist consists of 8-10 elements arranged in three rows . metacarpus includes five bones (according to the number of fingers). Each finger has three phalanges, with the exception of the first finger, which consists of two phalanges. Unlike the intercarpal articulation of reptiles and birds, the movable joint located in mammals between forearm and hand(as in amphibians).
hind limb mammals also has three divisions - thigh, leg and foot(Fig. 82 B). Femur massive and inserted with a rounded head into the acetabulum of the pelvic girdle. The tibia includes two bones - the greater tibia and the lesser tibia. The larger tibia is the inner element of the lower leg, and the fibula is its outer part.
Between thigh and lower leg joint, covered kneecap, which evolutionarily formed their ossified muscle tendons. The surfaces of the articulating bones are covered with cartilaginous formations. Ankle joint mammals is located between the lower ends of the tibia of the lower leg and the proximal foot.

Rice. 82. Pelvic girdle (A) and hind limb (B) of a fox:
1 – ilium, 2 - ischium, 3 - pubic bone, 4 - acetabulum, 5 - femur, 6 - patella, 7 - tibia, 8 - fibula, 9 - tarsus, 10 - metatarsus, 11 - phalanges, 12 - talus, 13 - calcaneus
Tarsal elements feet are arranged in three rows and form an outgrowth - calcaneus characteristic of mammals. Metatarsus consists of five bones, to which the phalanges of the fingers are attached. As in the forelimbs, there are three of them in each finger, except for the first (inner), which has only two phalanges.
Mammals are characterized by a special position of the limbs relative to the body: they are located under the body, which contributes to the development of greater maneuverability of movements. In this case, the elbow joint is directed backwards, and knee-joint, on the contrary, is oriented anteriorly.
Due to the diversity of mammals, their way of life and methods of movement, the relative length of the limbs, the ratio of the constituent elements, the configuration of the bones, the number of fingers, etc., vary significantly. For example, in bats, the length of the forelimb, especially the phalanges of the fingers, is sharply increased, serving to attach the flying membrane.
On the contrary, the forelimb of the mole is shortened, the palmar part of the hand is expanded, and the sixth finger is well developed. In whales, the hand is sharply elongated due to an increase in the number of phalanges of the fingers, as a result of which the limb acquired great resemblance with the fin of a lobe-finned fish. Significantly reduced and modified fingers of animals with large biomass and fast running (equids, artiodactyls).
One of the features of the mammalian skeleton is great development cartilage in young animals and its long-term preservation in the postembryonic period of development. This is due to the peculiarities of the growth of cartilage and bone in connection with the growth of the animal itself. It is the cartilage that grows with its entire mass (interstitially), growth occurs quickly and the proportions of the parts can be preserved, while the bone grows more slowly and only from the surface, so that the parts of the skeleton that have appeared always remain at the same distance and the preservation of the proportion of the parts during bone growth turns out to be impossible. That is why young animals always contain a lot of cartilage, and especially in places where growth is strong. So, long bones limbs always consist of the middle part - the diaphysis, which ossifies first of all, and the ends that remain cartilaginous for a long time - the epiphysis. Ossification of the diaphysis begins primarily from the periphery - perichondrally, i.e., due to cells originating from the connective tissue that dresses the cartilage - from the perichondrium; then the internal ossification of the cartilage joins it - endochondral ossification; only later, when the cavities of the bone marrow appear in the diaphysis by disintegration of the bones, the epiphyses begin to ossify endochondral; between the diaphysis and the epiphyses, however, cartilage remains for a long time, with the help of which the bone grows in length with the preservation of proportions (Fig. 530).
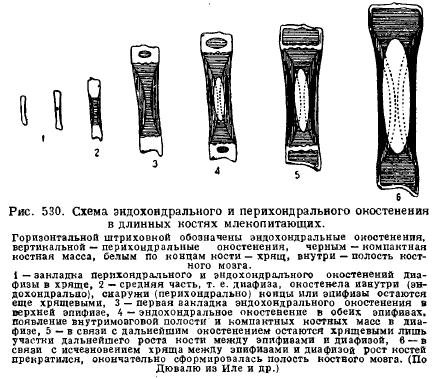
Spine mammals is characterized in contrast to Sauropsida by the absence of free articulations between the vertebrae, with the exception of the first and second vertebrae. Intervertebral elastic connective tissue discs lie between the vertebral bodies. In general, this causes less flexibility of the spine, which is due to the more specialized movement of mammals with the help of limbs.
Ossification of the vertebral bodies occurs through the cartilaginous epiphysis. The bodies are either flat or slightly convex, and are slightly opisthocoelous in most modern ungulates. On the vertebrae, the spinous processes are well developed, especially in the thoracic region, as well as the transverse processes, which sit on the arcs; at the base of the latter there are articular processes.

The number of cervical vertebrae is usually 7 (Fig. 531), regardless of the length of the neck, so that the length of the neck, in contrast to birds, depends solely on the length of the vertebrae. Only the manatee (Manatus) and one species of sloth (Choloepus hoffmani) have 6 cervical vertebrae, while the three-toed sloths (Bradypus) have 9. The neck of cetaceans, which has almost disappeared due to the aquatic lifestyle, still contains 7 flat, partially merging vertebrae, while in the neck of a giraffe these same 7 vertebrae are very long. The first 2 cervical vertebrae are modified into an atlas and epistrophy, connected by real joints, and in monotremes, the odontoid process still functions as a special bone, and has not fused, as in other mammals, with the body of the epistrophy. The rudimentary ribs of the cervical region fuse with the vertebrae, resulting in the formation of perforated transverse processes. Partial or complete fusion of the cervical vertebrae occurs in cetaceans; in jerboas, all cervical vertebrae grow together with the exception of the 7th, in the marsupial mole (Notoryctes) they merge from the 2nd to the 6th, in the mole - from the 3rd to the 4th.
The number of thoracic vertebrae, which are characterized by the attachment of ribs to them, varies from 9 [in one cetacean (Hyperoodon) and armadillo (Tatusia)] to 25 [in a sloth], but usually there are 12-13. On the thoracic vertebrae, processes are well developed: spinous (proc. spinosus), transverse, or diapophyses (proc. transversus), articular, or zygapophyses (proc. articularis); the ribs are articulated with their heads to the body of one or two adjacent vertebrae and tubercles to the transverse processes. They consist of two parts: a bone rib - proximal and cartilaginous - distal. Only the ribs of the anterior thoracic vertebrae reach the sternum, the posterior ribs terminate freely (false ribs). There are from 2 to 9 lumbar vertebrae, which are characterized by the absence of ribs, which are included here in the composition of the transverse processes, in mammals, usually 6-7.
The sacral vertebrae of mammals are those vertebrae that, fused together, articulate with ilium. There are 1-10 such vertebrae, but only 1-2 of them are true sacral vertebrae, that is, those in which articulation with the pelvis occurs through independent ossifications corresponding to the sacral ribs; the remaining sacral vertebrae are false, i.e., tail vertebrae adhering to the sacrum.
The number of vertebrae entering the caudal region varies enormously: from 3 ( the bats, gibbon) to 49 [lizard (Manis macrura)].
Sternum(sternum) of mammals develops from the fusion of the ventral ends of the ribs, which then form one unpaired plate, consisting of a number of sections (Fig. 532).
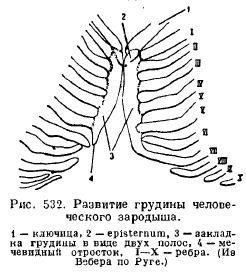
Connected to the clavicles (if any) and to the first pair of ribs, the expanded anterior sternum is called the handle (praesternum, manubrium); this is followed by a section called the body (mesosternum, corpus), often disintegrating into a series of bones sequentially located between successive pairs of ribs; even further back lies the so-called xiphoid process (xiphisternum). Not all ribs are in connection with the sternum, a more or less significant number of false ribs end freely (Fig. 533).

In cloacal mammals (Monotremata), due to the presence of coracoids absent in all other mammals, a cartilage of the presternum (prosternum) is laid in front of the handle of the sternum, which is in connection with the coracoids, which is then replaced by a developing bone - the episternum (episternmn), which brings the cloacal to reptiles.
Scull mammals a number of features differs from the skull of reptiles. However, these features did not appear immediately, but through a gradual change, which can be found by studying the skulls of fossil animal lizards (Theromorpha), especially from the order of animal-toothed (Theriodontia). The skull of mammals is a tropibasal skull, but with a shortened interorbital part, due to which it looks like a platybasal one, with a large cranium, with a clearly pronounced tendency for individual bones to fuse together,
The location of the bones in the skull of mammals can be judged from the attached diagram (Fig. 534).
On the lateral occipital bones, outward from the articular tubercles, there is usually an occipital process (proc. paroccipitalis), which is especially well developed in rodents and in many ungulates. Ahead of the articular tubercles, there is usually an opening for the exit of the hypoglossal (XII) nerve (for. condyloideum anterius).
Several ossifications are laid in the cartilage of the auditory capsule, which, however, very early merge into two bones: the one lying in front - the stony (petrosum) and the one lying outside and behind - the mastoid (mastoideum); both of these bones, fusing together, give a stony-mastoid, or simply a stony bone (petromastoideum, petrosum or perioticum), which encloses the internal auditory labyrinth. Perioticum borders in the posterior part on the main and lateral occipital bones, and in the anterior part on the sphenoid and wing-sphenoid bones; since vessels and nerves pass between these bones and the petrous bone, significant gaps remain here: behind - the posterior torn hole (for. lacerum posterius), or the jugular hole (for. jugulare) for the passage of the glossopharyngeal (IX) nerve, branches of the vagus ( X) nerve and for the accessory (XI) nerve, as well as for the internal jugular vein, and in front - the anterior torn hole (for. lacerum anterius), through which the internal carotid artery passes into the skull. Sometimes the connection of the petrous bone with the skull is so weak that it can easily be separated along with the tympanic bone that has adhered to it, or even completely falls out when the skull is macerated, as is the case with cetaceans.
Outside the ear region, two integumentary bones are formed. One of them, the origin of which is still not sufficiently clarified, develops in the region of the external auditory canal in the form of a semicircular small bone. This is a tympanic bone (tympanicum), which is probably a homologue of the angular bone (angulare) mandible reptiles, which passed into the parotid region along with other bones of the lower jaw due to changes that occurred in the jaw joint. Initially located in the form of a semicircular or annular bone in the region of the auditory canal, it often grows and forms, usually together with the stony bone and sometimes with the scaly, the external auditory meatus (meatus auditorius externus). The tympanic membrane is stretched over the tympanic bone. Deeper inward, the tympanic bone can also take part in the formation of the walls of the middle ear. Often, the tympanic bone on the underside of the auditory region swells into a thin-walled bubble (bulla ossea).
The second bone of skin origin, which takes part in the formation of the skeleton of the ear region, is the squamous bone (squamosum); it received exceptional development and importance in mammals in connection with the attachment of the lower jaw to it. In monkeys and humans, all these bones of the ear region fuse together, forming a complex temporal bone (temporale); the squamous bone here is only a part, namely the scales (squama temporalis) temporal bone, while the tympanic bone is the tympanic part (pars tympanicum), and the parotid cartilaginous bones proper form the stony (pars petrosa) and mastoid (pars mastoidea) parts. The degree of development of the squamous bone and its participation in the formation of the walls of the brain box and the auditory canal are very different, but the squamous bone always has an articular surface (cavitas glenoidea) for articulation of the lower jaw. Thus, in mammals, unlike all other vertebrates, there is a very special original articulation of the lower jaw: not to the quadrate bone, but to the squamous bone of the skull. The articular surface for the lower jaw lies on the base of the outward and forward zygomatic process(proc. zygomaticus). In monotremes above the articular surface, it is pierced by the temporal canal (can. temporalis), which is especially pronounced in the platypus. Behind the articular fossa there is a process (pros. postglenoideus).
In front of the auditory region, the base of the skull is formed by the main sphenoid bone (basisphenoidale), on inner surface which has a more or less significant depression - the so-called Turkish saddle (sella turtica), in which the pituitary gland is placed. On the sides of the main sphenoid bone, forming the lower parts of the anterior wall of the cranium, the wing-sphenoid bones (alisphenoidea), usually fused with it into one bone, form large wings (alae magnae) of this bone. The pterygo-sphenoid bones are equipped with two holes: oval (for. ovale) for the passage of the third branch of the trigeminal nerve (V3) and round (for. rotundum) for the passage of the second branch of the same nerve (V2). However, the first often merges with the anterior ragged foramen (for. lacerum anteris), and the second often merges with the wedge-orbital foramen (for. sphenorbitale), limited both by the pterygo-sphenoid bone and the orbital-sphenoid bone lying in front of it. Through this hole pass the nerves oculomotorius III, block-shaped (trochlearis IV), abducens (abducens VI) and the first branch of the trigeminal nerve.
The cranial cover here is covered by the parietal bones (parietale), which are often fused with each other (in monotremes, some marsupials and ungulates); between them and the upper occipital bone, two more bones are laid, merging into one interparietal bone (interparietale), which in some orders is preserved as an independent bone (cetaceans, marsupials, hyraxes, aardvarks, woolly wings and some rodents), while in others it fuses or with the parietal (in some rodents, sirens and ruminants), or with the upper occipital bone (in carnivores, one-hoofed animals, primates).
In front of the main sphenoid bone lies the anterior sphenoid bone (praesphenoideum) with a recess for the chiasm of the ophthalmic nerves; on the sides of it, usually growing together with it, are the orbital-sphenoid bones (orbitosphenoidea) with a hole for the passage of the orbital nerves (for. opticum), called in the case of fusion with the anterior and main sphenoid bones, small wings (alae parvae). As mentioned above, between both pterygoid bones on each side there are slit-like holes through which the nerves of the eye muscles and the first branch of the trigeminal nerve (for. sphenorbitale) pass.
On top of this row of bones correspond the integumentary frontal bones (frontale) - usually well-developed paired bones, sometimes fused together and bearing rods in hollow-horned ones, and stumps for antlers in deer. Covering the cranium, the frontal bones are limited from above and from the inside of the orbit, often forming processes: preorbital (pros. praeorbitalis and behind-orbital (pros. postorbitalis). Sometimes around the eyes it is formed due to fusion zygomatic bone(jugale) with a frontal bone, an orbital ring (some insectivorous and carnivorous, ruminant, one-hoofed, semi-monkeys). In monkeys, the process of separation of the eye socket from the temporal cavity goes so far that a bone wall is formed between the orbit and the temporal cavity.
In the anterior wall of the orbit lies a small integumentary lacrimal bone (lacrymale), perforated by the opening of the lacrimal canal (for. lacrymale).
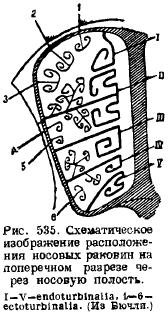
In the nasal region, several ossifications are laid in the cartilaginous nasal capsule. First, along the cartilaginous nasal septum, a bone septum (lamina perpendicularis) develops in the middle, corresponding to the middle olfactory bone (mesethmoideum); secondly, in back wall the nasal capsule and in the anterior wall of the brain cavity develop two lateral plates, perforated by numerous [only in the platypus these holes are replaced by one (for. olfactorium)] holes for the passage of the olfactory nerve - cribriform plates (lamina cribrosa), corresponding to the lateral ethmoid bones (ethmoidalia lateralia) . From the nasal septum, ossification spreads from the dorsal side along the sides, goes along the side walls of the nasal capsules and ends at its bottom, forming numerous protrusions inside the nasal cavity from the sides. These are olfactory shells (ethmoturbinalia), which, together with the lateral plates and with the cavities of the cellular structure, sometimes form a very complex nasal labyrinth. The location of the shells can be seen from the attached schematic drawing (Fig. 535). From the integumentary nasal bone (nasale), covering this area from above, the shells (nasoturbmalia) hang down inside the nasal cavity, also one on each side. Bottom on nasal septum an unpaired bone leans, which was previously considered a vomer (vomer). However, recent studies have shown that, in addition to this unpaired integumentary bone, two paired bones develop at the base of the skull, called the presomer (praevomer); and since in reptiles the vomer is in the form of a paired bone, and the unpaired bone in this region was the parasphenoid, then the mammalian vomer can be considered the homologue of this last bone of reptiles, and the pre-vomers can be considered vomers. In front, the walls of the nasal cavity are the jaw bones (maxillare), on which shells are formed that grow into the nasal cavity; these are the jaw shells (maxilloturbinale).
The front side of the skull ends in front with the premaxillary bones (praemaxillare). In anthropomorphic monkeys and in humans, this bone fuses very early with the jawbone, while in bats it is reduced to a small bone or even absent altogether. With its vertical process, the premaxilla limits the nasal opening in front and extends between the nasal bone and the jaw far back, sometimes to the frontal bone. From below, the intermaxillary bones take part in the formation of the hard palate. They limit the front and sides on the hard palate openings of the palatine-posterior (stenson) canal (for. incisiva), connecting oral cavity with a Jacobson organ. The processes of the premaxillary bones lying outside these palatonasal openings are superimposed for each bone independently, they are probably homologues of the paired vomer of reptiles (praevomer) and are preserved in the platypus as independent bones. The incisors are located on the intermaxillary bone.
The jaw bones (maxillare), as already mentioned, limit the bottom of the nasal cavity, forming a hard palate with horizontal processes. On the inner side of the nasal cavity, to its side walls, formed by the jaw bones, grows the lower wearable shell (maxilloturbinae), which is laid as an independent bone.
The continuation of the hard palate backwards are the horizontal processes of the palatine bones (palatinum), which limit the internal nasal passage as side walls. They stand vertically, and behind them are also adjoined vertically standing pterygoid bones (pterygoideum), which in humans grow together with the sphenoid bone, forming its pterygoid processes (proc. pterygoidei).
The anterior part of the skull undergoes significant modifications in connection with the transition to an aquatic lifestyle in cetaceans, sirens, and in connection with the development of the trunk in elephants.

In cetaceans (Fig. 536), the nostrils are pushed far back into the interorbital region and lead into a vertical canal, correspondingly opening through the choanae into the oral cavity; nasal bones lie behind the nostrils, leaning on the frontal bones, and almost adjacent to the interparietal bone; also, the jaw bones overlap the frontal bones, reaching almost to the parietal bones, which are strongly shifted to the sides by the interparietal bone.
Sirens have the same backward movement of the nostrils, but not so far; the nasal bones here are even more reduced and may even be absent (Fig. 537).

In elephants with an exorbitant development of tusks and powerful molars, and due to the presence of a trunk, the normal arrangement bones: the nostrils are strongly pushed back, in connection with this, the nasal canal takes an almost perpendicular position; the frontals are very short, the premaxillae are very large and extend far back; the jaw bones are shifted under the base of the skull; the entire skull becomes very massive with an extremely developed pneumatism of the bones (Fig. 538).

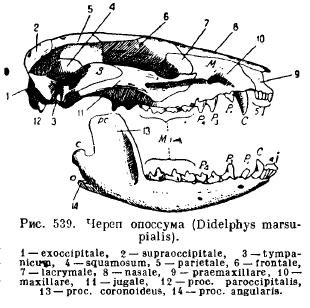
The lower jaw of mammals, unlike all other vertebrates, develops on each side from only one dentary bone (dentale); both halves either remain connected to each other by means of cartilage, a seam, or grow together (one-hoofed, elephant, bats, monkeys). Each half consists of a horizontal body (corpus) on which the teeth sit, and an ascending part with processes: the anterior coronoid process (pros. coronoideus), the articular process lying behind it (pros. articularis) and, in lower mammals, the angular process going backwards (pros .angularis); in most marsupials and in individual rodents, this process turns into a plate extending inward (Fig. 539).
Such a structure of the lower jaw and its peculiar attachment to the skull lead us to the question of the origin of these features in mammals. History individual development and paleontological data provide the answer to that question. In mammalian embryos, palatal-square and Meckel's cartilages are formed. However, the ossification of the palatine-square cartilage occurs without connection with the articulation of the lower jaw and does not lead to the formation of a square bone, as in reptiles, and will give a new auditory bone - an anvil (incus), while Meckel's cartilage, ossified in its posterior part, gives a non-articular bone (articulare), as in other maxillary, and the third auditory bone - malleus (malleus). The first auditory ossicle, stirrup (stapes), corresponds to the column of amphibians, reptiles and birds and, as we already know, is a modified suspension (hyomandibulare), i.e., the proximal hyoid arch. Like the column (columella auris), the stirrup covers the oval window of the auditory labyrinth, and is perforated, as in some reptiles, by a hole through which the facial artery (a. facialis) passes in some lower mammals (Fig. 540 and 541).

Thus, in mammals, the square bone turned into an anvil, the articular bone into a malleus; and since a small integumentary bone grows to the malleus, forming its middle process, it is believed that this latter corresponds to the integumentary articular bone (goniale) of reptiles. Lying next to this last bone, the integumentary angular bone of reptiles also passes into the ear region, becoming the tympanic bone (tympanicum). These changes are visible in the attached diagram (Fig. 541).
Paleontological data confirm the possibility of the above transformations. Among the animal-toothed animal lizards (Theriodontia) there are those in which the dentary is especially developed, reaching behind the place of articulation of the lower jaw with a square brush. At the same time, the squamosal descends along the quadrate to the same articulation. The dentary and squamosal bones thus come closer and closer.
Finally, in some animal-toothed animals, the dentary and squamosal bones take part in articulation along with the articular and quadrate bones.
As for the hyoid apparatus of mammals, it consists of a body, a modified basal part (copula) of the hyoid arch, anterior horns (cornua anteriora) formed by the lower part of the hyoid arch, and posterior horns (cornua posteriora) transformed from the first branchial arch. The anterior horns consist of several ossifications (up to four); the segment facing the skull often adheres to the petrous bone, forming the styloid process (proc. styloideus).
The second and third gill arches gave rise to the formation of the thyroid cartilage (cartilago thyreoidea.)
Limb belts. The shoulder girdle of mammals is characterized primarily by the reduction of the crow bones (procoracoideum, epicoracoideum, coracoideum), the tendency to the disappearance of the clavicles (clavicula) and a well-developed crest of the scapula (spina scapulae), continuing into the acromial process (acromion).
However, in monotremes, the shoulder girdle bears a strong resemblance to reptiles. The scapula is equipped with an acromial protrusion, to which the clavicle adjoins in front, and wide and short crow bones (coracoideum), going to the sternum, are articulated behind the scapula. In front of them, there are wide procoracoids (procoracoideum) in the form of independent bones. The clavicles (clavicula) are medially connected to the breastplate (episternum) (Fig. 542). In other mammals, cracoids are laid in the form of two ossifications, but they lose their connection with the sternum and adhere to the scapula, forming a coracoid process (proc. coracoideus); procoracoids disappear completely (Fig. 543), clavicles disappear in some carnivores and ungulates in connection with adaptation to fast running, and in cetaceans and sirens in connection with the transformation of the forelimbs into fins.

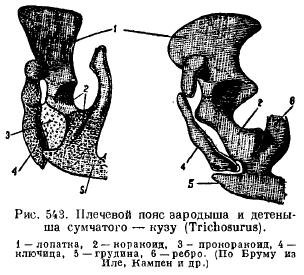
The pelvic girdle of mammals has a typical arrangement of elements when laying; its peculiarity is, firstly, the presence of an additional bone in the acetabulum (acetabulare), and secondly, the fusion of all these bones on each side into one nameless bone (innominatum) and the fusion of the pubic and ischial bones together, so that a closed joint is formed. all sides of the hole between these bones (for. obturatum) (Fig. 544). In monotremes and marsupials, a special bone is attached to the pubic bone below to support the bag (marsupiale).
limb skeleton in the original forms of mammals, it is due to their adaptation to fast movements - running with the help of four limbs, which are located under the body, so that the latter does not hang between the limbs, as in most reptiles, but rests directly on them. The ability to run is due to a different position of the limbs in mammals. It is in them that the limbs are oriented so that the elbow joints of the forelimbs are directed backward, and the knee joints of the hind limbs are directed forward. When moving, the forelimbs are thrown forward and attract the body by bending the carpal and elbow joints; the hind limbs push the body forward by straightening the leg at the knee joint.
According to the number of bones included in the limbs and their location, the skeleton of the limbs of the ancestral forms of mammals has a very typical structure of a primitive five-fingered limb and has undergone big changes within the class only in the process of adaptation to various kinds of movements in different conditions environment.
In the forelimb, the presence in the distal section is characteristic humerus holes (for. entepicondylare), the presence of a proximal protrusion of the ulna (olecranon), the presence of a pisiform bone (pisiforme) and the fusion of the fourth and fifth distal carpal bones (carpalia dis talia 4 + 6 \u003d hamatum, or uncinatum) and the central proximal with the intermedial (centrale dist + intermedium = lunatum, or semilunare). Due to the fact that the nomenclature of the bones of the mammalian wrist, adopted in human anatomy, is often used, we present a table of these names in a diagram indicating the location of the bones.
The hind limb of mammals is characterized primarily by the fact that it is turned at the knee joint in front of the thigh and has well-developed processes on the proximal part - trochanters, including 2-3 (trochanter major, minor and tertius); in the lower leg, the tibia is more developed, there is a bony patella (patella) that protects the knee joint. In the tarsus there are only two well-developed proximal bones: the inner talus (talus = astragalus), formed from the fusion of the intermediate and central proximal (intermedium + centrale proximale), and the outer calcaneus (calcaneus), corresponding to the fibular bone of the tarsus (fibulare). From the bones of the middle row, the central distal (centrale distale) turned into a boat-shaped (naviculare = scaphoideum). The tibial bone was either reduced or accreted to this latter.
The layout of the bones of the tarsus and metatarsus and the synonymy of the names are as follows:
In connection with the adaptation to movement in various environments, the limbs of mammals have undergone very significant changes, and these changes can be traced especially well on the skeleton. Unfortunately, we do not have paleontological data on the structure of the limbs of the ancestors of mammals. There is no doubt that the first mammals had little specialized limbs of the type described above. It is precisely these limbs that the most primitive marsupials of modern mammals have, for example, opossums - marsupial rats (Didelphys), and insectivores leading an arboreal lifestyle. However, it would be wrong to conclude from this that the ancestors of mammals were certainly inhabitants of forests and, moreover, forms that climbed trees. In addition, the limb of the opossum, although it serves to move through the knots of trees, is by no means a typical specialized arboreal limb, but retains its universal five-fingered type. Such limbs could best be preserved in animals that not only ran on the ground, but also climbed trees, as well as animals that climbed mountains. The specialization of the limbs of mammals proceeded simultaneously with the differentiation of orders and other smaller systematic units, and convergence took place.
Limbs undergo especially profound modifications when adapted for flight, for swimming in water, for movement underground; slow movements on the ground and an arboreal way of life do not stand in connection with deep specialization.
In bats (Chiroptera), the forelimb is turned into a wing, and the flying membrane is stretched between four (2-b) strongly elongated fingers, devoid of claws; the first finger is short and has a strongly developed claw at the end; this finger does not play any role in flying, it is used for climbing; in the wrist, the navicular, lunate and central bones fuse into one; the forearm is elongated and consists of a well-developed radius, while the ulna is reduced and adheres to the radius. The flying membrane also captures the hind legs. Between the latter, it is developed differently in bats (Fig. 545).
In water-swimming mammals, the limbs are transformed into fins in a similar way, as we have already seen in reptiles. In this case, the limbs of whales and sirens have changed especially strongly. In contrast to the wing, the forelimb is significantly shortened in its proximal part. In whales, the bones of the shoulder and forearm have turned into short and wide plates, giving a place for attaching powerful muscles. All parts of the limb are fixedly connected to each other, forming a strong paddle. In this case, the number of phalanges can be greatly increased. We have seen the same changes in reptiles. The increase in the number of phalanges depends on the fact that the diaphyses of the phalanges do not fuse with the epiphyses and ossify from separate centers. This separate ossification stands in connection with the slow process of ossification in these animals. The slowness of the ossification process leads to the fact that most of the bones at their junctions remain cartilaginous.
The fins of sirens (Sirenia) have an external resemblance to those of cetaceans: the shoulder and forearm are articulated with each other, so that they can bend; there are almost all the bones of the wrist and all five fingers with a slightly changed number of phalanges (Fig. 546).
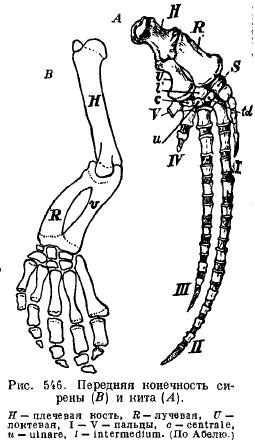
The hind limbs modern whales and sirens are reduced so much that only the remains of the pelvis remain from them (Fig. 547 and 548).
Another type of adaptation of limbs to movement in the pod we see in pinnipeds (Pinnipedia). Here the tail almost disappears, and the hind legs move back, playing the role motor organ. According to the degree of development of adaptation to movement in water, the three families of this order represent, as it were, three stages (not genetically). In walruses and in eared seals (Otariidae), the hind limbs can still be bent forward under the body, but in walruses, the structure of the limbs is more primitive, they are shorter and more mobile. In seals (Phocidae), the hind legs can no longer bend forward under the body, but claws are still preserved on the toes (fins).
In burrowing animals, the limbs also undergo very strong modifications: the limbs of the marsupial mole (Notoryctes typhlops) have changed especially strongly. It has two ridges on the shoulder blade, like many burrowing animals, which is due to the strong development of the triceps muscle of the shoulder; there is a clavicle, like all burrowing mammals. There is a very strong ridge on the humerus, branching in a fork-like manner and serving to attach strong (necessary for digging) deltoid and chest muscle; the internal condyle of the shoulder is strongly developed due to strong development muscles of the flexors of the ulna (proc. olecranon), which serves as the site of attachment of the triceps muscle; the wrist is peculiarly changed, becoming very strong; The 1st and 2nd fingers are moved to the palmar side, and only the 3rd and 4th are used for digging, equipped with huge claws, especially the 3rd; the number of phalanges in the fingers is very small: 2 and 1. 5th metacarpale with a wide flat claw, which increases the burrowing surface; in the plane of the palm there is a sesamoid bone, to which a muscle is attached that flexes the fingers, which plays an important role in digging; it increases the strength of the wrist; like other burrowing forms, symphysis pubic bones very short; the ischium fuses with the transverse processes of the last two sacral vertebrae; on the thigh, knee and tibia, crests and outgrowths are strongly developed - places of attachment of muscles; the tibia and fibula (tibia and fibula) are firmly fused, as in the golden mole (Chrysochloris); the sole is turned outward, as in Chrysochloris; The 5th toe is bent onto the plantar surface, the 5th nail on the foot is wide and spatulate, the 4th is long and crescent-shaped, the 3rd and 2nd become shorter and wider; on the sternum there is a strongly developed crest, as in the golden mole, mole rat (Spalax) and mole (Talpa) (Fig. 549).
The strong development of ridges on the bones for the attachment of muscles, the shortening of the carpus, the strong connection of the carpalia, the reduction of some phalanges and the strengthening of others that play a role in digging - all these features are characteristic of burrowing mammals in general. Ho, depending on the soil in which the animal digs, differences will be observed. When digging in soft soil (for example, in a mole - Talpa) (Fig. 550), the brush expands greatly, it acts like a shovel and has a tendency to develop the 6th finger. The forelimb of the mole is comparable to a shovel used for digging in soft ground. On the other hand, in those forms that dig in hard ground, the hand becomes narrower, only a few fingers take part in digging, for the most part the 3rd and 4th, developing more strongly than others, as in Notoryctes and Chrysochloris. This brush is comparable to a spade for digging in hard ground.
![]()
We see the same thing in mammals that tear apart the hard constructions of termites, for example, in the anteater (Myrmecophagus), in the tamandua (Tamandua). These animals walk, leaning on the outer edge of the feet, bending their claws.
Adaptations of mammals to rapid movement on the ground caused very significant changes in the limbs. Movement on the ground, which was characteristic of primitive mammals, was carried out by relying on the entire hand and foot; original forms mammals were primarily plantigrade or plantigrade, i.e., they leaned on the entire foot when walking, like, for example, bears. But such a structure allows only relatively slow walking in comparison with swift-footed antelopes or horses, or jumping hares, kangaroos, jerboas. During phylogenetic development, we see a gradual rise above the ground of the carpus and tarsus (carpus and tarsus), followed by the metacarpus and metatarsus, but the phalanges still touched the ground entirely. In the end, the phalanges also straightened out, the front and hind limbs began to rest on the ends of the fingers. We can now trace the stages of this evolution step by step not only in ungulates, but also in predatory mammals that have acquired the ability to run fast. As the limbs straighten out, the extreme toes no longer touch the ground, cease to function, and undergo atrophy. This results in a smaller contact surface with the ground. Changes in the distal part of the limbs entailed changes in the proximal part: the carpalia and tarsalia are located more and more closely next to each other, the ulna and fibula are reduced, the limbs begin to move only in the sagittal plane, the median metapodia merge into one bone.
Two developmental pathways lead to the formation of the limbs of modern ungulates. In one row, the axis of the limb passes in the middle of the third finger, and the latter develops most strongly, while the rest become rudimentary. These are equids (Perissodactyla), mesaxial (Mesaxonia). In others, the axis passes between the third and fourth fingers, which develop evenly: artiodactyls (Artiodactyla), paraxial (Paraxonia). Lateral fingers are reduced. The development of equids is most clearly seen in the phylogenetic development of the horse's limbs (Fig. 551). The development of the artiodactyl state was due, among other things, to the fact that it was preceded by a reduction of the first toe, so that in the transition from planting it almost did not take any part, while in the ancestors of one-hoofed animals the development of the fifth toe was initially normal,
Interestingly, both artiodactyls and equids arose repeatedly in the evolutionary process and in some cases led to a very specialized type, for example, in the South American Proterotlieriidae (extinct).
The surviving toes are clad with hooves, which may further differentiate depending on lifestyle and habitat. Thus, the goats, holding on to the rocks, have hooves that are steep and narrow, and provided with a sharp edge; at reindeer they are very long, widely forked and serve as a kind of skis that prevent the animal from falling into snow and swamps.
Simultaneously with the development of walking on the fingers, there is a change in the wrist (carpus) and tarsus (tarsus). It goes differently in artiodactyls and equids due to a different distribution of gravity. Finally, in connection with the transition to natal walking, the importance of the ulna (ulna) as a support for the shoulder gradually decreases: it decreases or merges with the radius (radius). In the same way, the fibula (fibula) is reduced and finally disappears in the hind limbs.
The significance and nature of the described changes are clearly visible from a comparison of the limbs of a camel and an elephant (Fig. 552). In the elephant, the hand (foot) remains short, while the shoulder and forearm (thigh and lower leg) are elongated. Conversely, the fused metapodia lengthen in the camel; the upper arm and forearm (as well as the thigh and lower leg) remain short. Due to the loss of a number of bones, the skeleton of the limb is simplified, the latter becomes lighter. The reduction of the fingers leads to less friction, the risk of damage, rupture of the ligaments is reduced. The intergrowth of the metapodia creates greater strength. The limb of an elephant is much heavier. The abundance of bones in the hand requires the connection of all fingers to obtain stability common skin. The limb of an elephant is a column of great strength, but devoid of the lightness that the limb of a camel has.
Another way to move quickly on the ground is to jump. This adaptation arose repeatedly in the history of mammals and, moreover, in a wide variety of phylogenetic trunks: among marsupials - in kangaroos (Macropus), among insectivores - in jumpers (Macroscelidae), and, finally, in different rows development of rodents: family. hares (Leporidae) from Duplicidentata; in the series of porcupines (Hystricomorplia), a group of striders (Pedetinae) developed; among the murines (Myomorpha) we have the Australian jerboa (Conilurus), the Old World jerboas (Jaculus, Alactaga), the American jumpers (Zapus) and the pouched jumpers (Dipodomys).
The development of the ability to jump is associated with the lengthening of the hind limb, namely the bones of the metatarsus (metatarsus). In addition, this results in the loss of extra extreme fingers, and the bones of the tarsus are interconnected very firmly. Examples are the feet of the ground hare (Alactaga) and true jerboas (Dipus).
The front legs of Alactaga are small and short. The earth hare uses them when digging, but does not touch the ground with them when jumping. Jumping is carried out only with the help of the hind legs and even without the help of the tail. The latter touches the ground only in a calm position, when the animal leans on its tail. The skeleton of the leg has the feature that the metatarsal bones of the 2nd, 3rd and 4th fingers merge into one bone. The ends of the metatarsal bones are not fused below, and the median (3rd finger) protrudes further forward. Of the phalanges connected to them, again, the middle ones are the longest. The 1st and 5th fingers are vestigial and do not touch the ground. Both of these fingers are the same length due to the fact that the metatarsal bone thumb longer than the 5th. The first has 2, the second - 3 phalanges.
In Dipus, specialization has gone even further: the 1st finger has disappeared without a trace, only a rudiment of the 5th remains. Middle finger twice weaker than the two extreme ones, and only slightly longer than them. The fingers are parallel. The middle finger is reduced because its function is taken over by the 2nd and 4th, at the ends of which there is emphasis when sitting and jumping. In further evolution, the 3rd finger must disappear: we have an unfinished type of device in Dipus.